
Siboglinidae is a family of polychaete annelid worms whose members made up the former phyla Pogonophora and Vestimentifera. The family is composed of around 100 species of vermiform creatures which live in thin tubes buried in sediment (Pogonophora) or in tubes attached to hard substratum (Vestimentifera) at ocean depths ranging from 100 to 10,000 m. They can also be found in association with hydrothermal vents, methane seeps, sunken plant material, and whale carcasses.

Any worm that lives in a marine environment is considered a water worm. Marine worms are found in several different phyla, including the Platyhelminthes, Nematoda, Annelida, Chaetognatha, Hemichordata, and Phoronida. For a list of marine animals that have been called "sea worms", see sea worm.

Polychaeta is a paraphyletic class of generally marine annelid worms, commonly called bristle worms or polychaetes. Each body segment has a pair of fleshy protrusions called parapodia that bear many bristles, called chaetae, which are made of chitin. More than 10,000 species are described in this class. Common representatives include the lugworm and the sandworm or clam worm Alitta.

In biochemistry, chemosynthesis is the biological conversion of one or more carbon-containing molecules and nutrients into organic matter using the oxidation of inorganic compounds or ferrous ions as a source of energy, rather than sunlight, as in photosynthesis. Chemoautotrophs, organisms that obtain carbon from carbon dioxide through chemosynthesis, are phylogenetically diverse. Groups that include conspicuous or biogeochemically important taxa include the sulfur-oxidizing Gammaproteobacteria, the Campylobacterota, the Aquificota, the methanogenic archaea, and the neutrophilic iron-oxidizing bacteria.
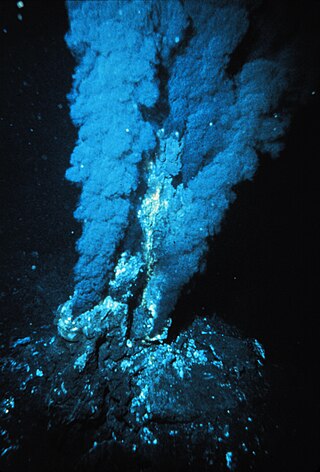
Hydrothermal vents are fissures on the seabed from which geothermally heated water discharges. They are commonly found near volcanically active places, areas where tectonic plates are moving apart at mid-ocean ridges, ocean basins, and hotspots. The dispersal of hydrothermal fluids throughout the global ocean at active vent sites creates hydrothermal plumes. Hydrothermal deposits are rocks and mineral ore deposits formed by the action of hydrothermal vents.

The Alvinellidae are a family of small, deep-sea polychaete worms endemic to hydrothermal vents in the Pacific Ocean. Belonging to the order Terebellida, the family contains two genera, Alvinella and Paralvinella; the former genus contains two valid species and the latter eight. Members of the family are termed alvinellids.

Riftia pachyptila, commonly known as the giant tube worm and less commonly known as the giant beardworm, is a marine invertebrate in the phylum Annelida related to tube worms commonly found in the intertidal and pelagic zones. R. pachyptila lives on the floor of the Pacific Ocean near hydrothermal vents. The vents provide a natural ambient temperature in their environment ranging from 2 to 30 °C, and this organism can tolerate extremely high hydrogen sulfide levels. These worms can reach a length of 3 m, and their tubular bodies have a diameter of 4 cm (1.6 in).

Mirocaris is a genus of shrimp associated with hydrothermal vents. Sometimes considered the only genus of the family Mirocarididae, Mirocaris is usually placed in the broader family Alvinocarididae. Mirocaris is characterized by a dorsoventrally flattened, non-dentate rostrum, as well as the possession of episodes on the third maxilliped through to the fourth pteropod. The genus contains two species, M. fortunata and M. indica. The two species are found in different oceans, and can be distinguished by the pattern of setation on the claw of the first pereiopod.
Paralvinella sulfincola, also known as the Sulfide worm, is a species of polychaete worm of the Alvinellidae family that thrives on undersea hot-water vents. It dwells within tubes in waters surrounding hydrothermal vents, in close proximity to super-heated fluids reaching over 300 °C (572 °F). The upper thermal limit for this polychaete is unknown; however, it is unlikely they can survive in constant temperatures over 50 °C (122 °F). It may tentatively be named a metazoan extremophile or, more specifically, a thermophile.

Lamellibrachia is a genus of tube worms related to the giant tube worm, Riftia pachyptila. They live at deep-sea cold seeps where hydrocarbons leak out of the seafloor, and are entirely reliant on internal, sulfide-oxidizing bacterial symbionts for their nutrition. The symbionts, gammaproteobacteria, require sulfide and inorganic carbon. The tube worms extract dissolved oxygen and hydrogen sulfide from the sea water with the crown of plumes. Species living near seeps can also obtain sulfide through their "roots", posterior extensions of their body and tube. Several sorts of hemoglobin are present in the blood and coelomic fluid to bind to the different components and transport them to the symbionts.

Terebellida make up an order of the Polychaeta class, commonly referred to as "bristle worms". Together with the Sabellida, the Spionida and some enigmatic families of unclear taxonomic relationship, they make up the subclass Canalipalpata, one of the three main clades of polychaetes. Like most polychaetes, almost all members of the Terebellida are marine organisms. Most are small, sessile detritivores which live in small tubes they build from mud or similar substrate, or burrow in the sand. Their central nervous system displays characteristic apomorphies.

Chrysomallon squamiferum, commonly known as the scaly-foot gastropod, scaly-foot snail, sea pangolin, or volcano snail is a species of deep-sea hydrothermal-vent snail, a marine gastropod mollusc in the family Peltospiridae. This vent-endemic gastropod is known only from deep-sea hydrothermal vents in the Indian Ocean, where it has been found at depths of about 2,400–2,900 m (1.5–1.8 mi). C. squamiferum differs greatly from other deep-sea gastropods, even the closely related neomphalines. In 2019, it was declared endangered on the IUCN Red List, the first species to be listed as such due to risks from deep-sea mining of its vent habitat.

Colleen Marie Cavanaugh is an American academic microbiologist best known for her studies of hydrothermal vent ecosystems. As of 2002, she is the Edward C. Jeffrey Professor of Biology in the Department of Organismic and Evolutionary Biology at Harvard University and is affiliated with the Marine Biological Laboratory and the Woods Hole Oceanographic Institution. Cavanaugh was the first to propose that the deep-sea giant tube worm, Riftia pachyptila, obtains its food from bacteria living within its cells, an insight which she had as a graduate student at Harvard. Significantly, she made the connection that these chemoautotrophic bacteria were able to play this role through their use of chemosynthesis, the biological oxidation of inorganic compounds to synthesize organic matter from very simple carbon-containing molecules, thus allowing organisms such as the bacteria to exist in deep ocean without sunlight.

Serpula is a genus of sessile, marine annelid tube worms that belongs to the family Serpulidae. Serpulid worms are very similar to tube worms of the closely related sabellid family, except that the former possess a cartilaginous operculum that occludes the entrance to their protective tube after the animal has withdrawn into it. The most distinctive feature of worms of the genus Serpula is their colorful fan-shaped "crown". The crown, used by these animals for respiration and alimentation, is the structure that is most commonly seen by scuba divers and other casual observers.
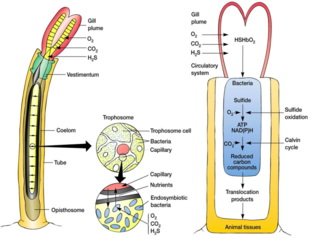
A trophosome is a highly vascularised organ found in some animals that houses symbiotic bacteria that provide food for their host. Trophosomes are contained by the coelom of the vestimentiferan tube worms and in the body of symbiotic flatworms of the genus Paracatenula.
Nautilia profundicola is a Gram-negative chemolithoautotrophic bacterium found around hydrothermal vents in the deep ocean. It was first discovered in 1999 on the East Pacific Rise at depth of 2,500 metres (8,200 ft), on the surface of the polychaete worm Alvinella pompejana. Nautilia profundicola lives symbiotically on the dorsal hairs of A. pompejana but they may also form biofilms and live independently on the walls of hydrothermal vents. The ability of N. profundicola to survive in an anaerobic environment rich in sulfur, H2 and CO2 of varying temperature makes it a useful organism to study, as these are the conditions that are theorized to have prevailed around the time of the earliest life on earth.
Vibrio diabolicus is a polysaccharide-secreting bacterium isolated from a deep-sea hydrothermal vent polychaete annelid, Alvinella pompejana. It is facultatively anaerobic, heterotrophic, and mesophilic.
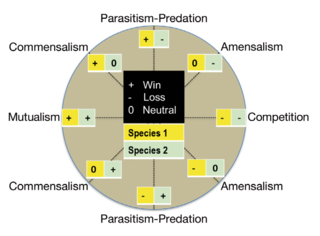
Microbial symbiosis in marine animals was not discovered until 1981. In the time following, symbiotic relationships between marine invertebrates and chemoautotrophic bacteria have been found in a variety of ecosystems, ranging from shallow coastal waters to deep-sea hydrothermal vents. Symbiosis is a way for marine organisms to find creative ways to survive in a very dynamic environment. They are different in relation to how dependent the organisms are on each other or how they are associated. It is also considered a selective force behind evolution in some scientific aspects. The symbiotic relationships of organisms has the ability to change behavior, morphology and metabolic pathways. With increased recognition and research, new terminology also arises, such as holobiont, which the relationship between a host and its symbionts as one grouping. Many scientists will look at the hologenome, which is the combined genetic information of the host and its symbionts. These terms are more commonly used to describe microbial symbionts.

Tevnia is a genus of giant tube worm in the family Siboglinidae, with only one species, Tevnia jerichonana, living in a unique deep-sea environment. These deep sea marine species survive in environments like hydrothermal vents. These vents give off gas and toxic chemicals with the addition of having superheated temperatures. The giant tube worm prefers environments such as these despite the harsh temperature and toxic sea water.

The hydrothermal vent microbial community includes all unicellular organisms that live and reproduce in a chemically distinct area around hydrothermal vents. These include organisms in the microbial mat, free floating cells, or bacteria in an endosymbiotic relationship with animals. Chemolithoautotrophic bacteria derive nutrients and energy from the geological activity at Hydrothermal vents to fix carbon into organic forms. Viruses are also a part of the hydrothermal vent microbial community and their influence on the microbial ecology in these ecosystems is a burgeoning field of research.
