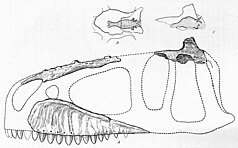
Spinosaurus is a genus of spinosaurid dinosaur that lived in what now is North Africa during the Cenomanian to upper Turonian stages of the Late Cretaceous period, about 99 to 93.5 million years ago. The genus was known first from Egyptian remains discovered in 1912 and described by German palaeontologist Ernst Stromer in 1915. The original remains were destroyed in World War II, but additional material came to light in the early 21st century. It is unclear whether one or two species are represented in the fossils reported in the scientific literature. The best known species is S. aegyptiacus from Egypt, although a potential second species, S. maroccanus, has been recovered from Morocco. The contemporary spinosaurid genus Sigilmassasaurus has also been synonymized by some authors with S. aegyptiacus, though other researchers propose it to be a distinct taxon. Another possible junior synonym is Oxalaia from the Alcântara Formation in Brazil.

Irritator is a genus of spinosaurid dinosaur that lived in what is now Brazil during the Albian stage of the Early Cretaceous Period, about 113 to 110 million years ago. It is known from a nearly complete skull found in the Romualdo Formation of the Araripe Basin. Fossil dealers had acquired this skull and sold it to the State Museum of Natural History Stuttgart. In 1996, the specimen became the holotype of the type species Irritator challengeri. The genus name comes from the word "irritation", reflecting the feelings of paleontologists who found the skull had been heavily damaged and altered by the collectors. The species name is a homage to the fictional character Professor Challenger from Arthur Conan Doyle's novels.

Giganotosaurus is a genus of theropod dinosaur that lived in what is now Argentina, during the early Cenomanian age of the Late Cretaceous period, approximately 99.6 to 95 million years ago. The holotype specimen was discovered in the Candeleros Formation of Patagonia in 1993 and is almost 70% complete. The animal was named Giganotosaurus carolinii in 1995; the genus name translates to "giant southern lizard", and the specific name honors the discoverer, Ruben Carolini. A dentary bone, a tooth, and some tracks, discovered before the holotype, were later assigned to this animal. The genus attracted much interest and became part of a scientific debate about the maximum sizes of theropod dinosaurs.

Acrocanthosaurus is a genus of carcharodontosaurid dinosaur that existed in what is now North America during the Aptian and early Albian stages of the Early Cretaceous, from 113 to 110 million years ago. Like most dinosaur genera, Acrocanthosaurus contains only a single species, A. atokensis. It had a continent-wide range, with fossil remains known from the U.S. states of Oklahoma, Texas, and Wyoming in the west, and Maryland in the east.

Suchomimus is a species of spinosaur dinosaur that lived between 125 and 112 million years ago in what is now Niger, north Africa, during the Aptian to early Albian stages of the Early Cretaceous period. It was named and described by paleontologist Paul Sereno and colleagues in 1998, based on a partial skeleton from the Elrhaz Formation. Suchomimus's long and shallow skull, similar to that of a crocodile, earns it its generic name, while the specific name Suchomimus tenerensis alludes to the locality of its first remains, the Ténéré Dese.

Ernst Freiherr Stromer von Reichenbach was a German paleontologist best remembered for his expedition to Egypt, during which the discovery of the first known remains of Spinosaurus was made.

Tyrannotitan is a genus of large bipedal carnivorous dinosaur of the carcharodontosaurid family from the Aptian stage of the early Cretaceous period, discovered in Argentina. It is closely related to other giant predators like Carcharodontosaurus and especially Giganotosaurus as well as Mapusaurus.

Spinosauridae is a clade or family of tetanuran theropod dinosaurs comprising ten to seventeen known genera. Spinosaurid fossils have been recovered worldwide, including Africa, Europe, South America and Asia. Their remains have generally been attributed to the Early to Mid Cretaceous.

Carcharodontosauridae is a group of carnivorous theropod dinosaurs. In 1931, Ernst Stromer named Carcharodontosauridae as a family, which, in modern paleontology, indicates a clade within Carnosauria. Carcharodontosaurids include some of the largest land predators ever known: Giganotosaurus, Mapusaurus, Carcharodontosaurus, and Tyrannotitan all rivaled Tyrannosaurus in size. Estimates give a maximum weight of 8–10 metric tons for the largest carcharodontosaurids, while the smallest carcharodontosaurids were estimated to have weighed at least 500 kilograms (1,100 lb).

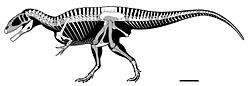
Rugops is a monospecific genus of basal abelisaurid theropod dinosaur from Niger that lived during the Late Cretaceous period in what is now the Echkar Formation. The type and only species, Rugops primus, is known only from a partial skull. It was named and described in 2004 by Paul Sereno, Jeffery Wilson and Jack Conrad. Rugops has an estimated length of 4.4–5.3 metres and weight of 410 kilograms. The top of its skull bears several pits which correlates with overlaying scale and the front of the snout would have had an armour-like dermis.

Bahariasaurus is an enigmatic genus of large theropod dinosaur. Bahariasaurus is known to have included at least 1 species, Bahariasaurus ingens, which was found in North African rock layers dating to the Cenomanian and Turonian ages of the Late Cretaceous. The only fossils confidently assigned to Bahariasaurus were found in the Bahariya Formation of the Bahariya oasis in Egypt by Ernst Stromer but were destroyed during a World War II bombing raid with the same raid taking out the holotype of Spinosaurus and Aegyptosaurus among other animals found in the Bahariya Formation. While there have been more fossils assigned to the genus such as some from the Farak Formation of Niger, these remains are referred to with much less certainty. Bahariasaurus is, by most estimations, one of the largest theropods, approaching the height and length of other large bodied theropods such as Tyrannosaurus rex and the contemporaneous Carcharodontosaurus. The aforementioned estimations tend to put it at around 11–12.2 metres in length and 4-4.8 tonnes in overall weight.

Deltadromeus is a genus of theropod dinosaur from the Aoufous Formation of Morocco.

Rebbachisaurus is a genus of sauropod dinosaur of the superfamily Diplodocoidea, that lived during the Late Cretaceous period in Africa and possibly also South America about 99-97 million years ago. Remains attributed to Rebbachisaurus have been found in Morocco, Niger, Algeria, Tunisia and possibly also Argentina, although only the Moroccan remains can be referred to the genus without doubt. The discovery of Rayososaurus, a South American sauropod nearly identical to Rebbachisaurus which may have actually have been the same animal as Rebbachisaurus, supports the theory that there was still a land connection between Africa and South America during the Early Cretaceous, long after it was commonly thought the two continents had separated.

Cristatusaurus is a genus of theropod dinosaur that lived during the Early Cretaceous Period of what is now Niger, 112 million years ago. It was a baryonychine member of the Spinosauridae, a group of large bipedal carnivores with well-built forelimbs and elongated, crocodile-like skulls. The type species Cristatusaurus lapparenti was named in 1998 by scientists Philippe Taquet and Dale Russell, on the basis of jaw bones and some vertebrae. Two claw fossils were also later assigned to Cristatusaurus. The animal's generic name, which means "crested reptile", alludes to a sagittal crest on top of its snout; while the specific name is in honor of the French paleontologist Albert-Félix de Lapparent. Cristatusaurus is known from the Albian to Aptian Elrhaz Formation, where it would have coexisted with sauropod and iguanodontian dinosaurs, other theropods, and various crocodylomorphs.

Mapusaurus was a giant carcharodontosaurid carnosaurian dinosaur from the early Late Cretaceous, approximately 93.9 to 89.6 million years ago, of what is now Argentina.

Sigilmassasaurus is a controversial genus of spinosaurid dinosaur that lived approximately 100 to 94 million years ago during the Late Cretaceous Period in what is now northern Africa. Named in 1996 by Canadian paleontologist Dale Russell, it contains a single species, Sigilmassasaurus brevicollis. The identity of the genus has been debated by scientists, with some considering its fossils to represent material from the closely related species Spinosaurus aegyptiacus, while others have classified it as a separate taxon, forming the clade Spinosaurini with Spinosaurus as its sister taxon.
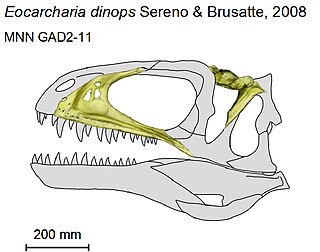
Eocarcharia is a genus of carcharodontosaurid theropod dinosaur from the Early Cretaceous Elrhaz Formation that lived in the Sahara 112 million years ago, in what today is the country of Niger. It was discovered in 2000 on an expedition led by University of Chicago paleontologist Paul Sereno. The type and only species is Eocarcharia dinops. Its teeth were shaped like blades and were used for disabling live prey and ripping apart body parts. Eocarcharia’s brow is swollen into a massive band of bone, giving it a menacing glare. It may have reached lengths of 6–8 m (19.7–26.2 ft).

The Kem Kem Group is a geological group in the Kem Kem region of eastern Morocco, whose strata date back to the Cenomanian stage of the Late Cretaceous. Its strata are subdivided into two geological formations, with the lower Ifezouane Formation and the upper Aoufous Formation, with the Gara Sbaa Formation and Douira Formation used in the southern Tafilalt region. It is exposed on an escarpment along the Algeria–Morocco border.

Oxalaia is a genus of spinosaurid dinosaur that lived in what is now the Northeast Region of Brazil during the Cenomanian stage of the Late Cretaceous period, sometime between 100.5 and 93.9 million years ago. Its only known fossils were found in 1999 on Cajual Island in the rocks of the Alcântara Formation, which is known for its abundance of fragmentary, isolated fossil specimens. The remains of Oxalaia were described in 2011 by Brazilian palaeontologist Alexander Kellner and colleagues, who assigned the specimens to a new genus containing one species, Oxalaia quilombensis. The species name refers to the Brazilian quilombo settlements. Oxalaia quilombensis is the eighth officially named theropod species from Brazil and the largest carnivorous dinosaur discovered there. It is closely related to the African genus Spinosaurus, and/or may be a junior synonym of this taxon.
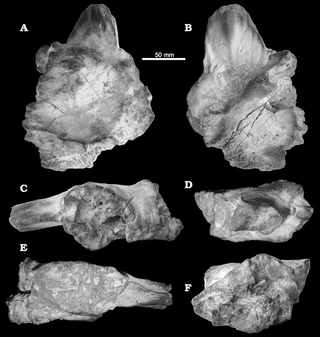
Sauroniops is a controversial genus of carnivorous basal carcharodontosaurid theropod dinosaur known from the Late Cretaceous Gara Sbaa Formation, and possibly also the Kem Kem Formation, both of Morocco. The type, and currently only, species is S. platytholus.