
Giganotosaurus is a genus of theropod dinosaur that lived in what is now Argentina, during the early Cenomanian age of the Late Cretaceous period, approximately 99.6 to 95 million years ago. The holotype specimen was discovered in the Candeleros Formation of Patagonia in 1993 and is almost 70% complete. The animal was named Giganotosaurus carolinii in 1995; the genus name translates to "giant southern lizard", and the specific name honors the discoverer, Ruben Carolini. A dentary bone, a tooth, and some tracks, discovered before the holotype, were later assigned to this animal. The genus attracted much interest and became part of a scientific debate about the maximum sizes of theropod dinosaurs.
Cryolophosaurus is a genus of large theropod dinosaur known from only a single species Cryolophosaurus ellioti, from the early Jurassic of Antarctica. It was one of the largest theropods of the Early Jurassic, with the subadult being estimated to have reached 6–7 metres (20–23 ft) long and weighed 350–465 kilograms (772–1,025 lb).
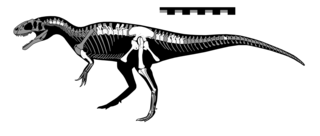
Gasosaurus is a genus of tetanuran theropod that lived approximately 171.6 to 161.2 million years ago during the middle of the Jurassic Period. The name "Gasosaurus" is derived from the English "gasoline" and the Greek σαῦρος. Only one species is currently recognised, G. constructus, from which the specific name honours the gasoline company that found the Dashanpu fossil quarry in Sichuan Province, China, now named as the Lower Shaximiao Formation.

Neovenator is a genus of carcharodontosaurian theropod dinosaur. It is known from several skeletons found in the Early Cretaceous (Hauterivian-Barremian) Wessex Formation on the south coast of the Isle of Wight, southern England. It is one of the best known theropod dinosaurs from the Early Cretaceous of Europe.

Allosauridae is a family of medium to large bipedal, carnivorous allosauroid theropod dinosaurs from the Late Jurassic. Allosauridae is a fairly old taxonomic group, having been first named by the American paleontologist Othniel Charles Marsh in 1878. Allosaurids are characterized by an astragalus with a restriction of the ascending process to the lateral part of the bone, a larger medial than lateral condyle, and a horizontal groove across the face of the condyles.

Eustreptospondylus is a genus of megalosaurid theropod dinosaur, from the Oxfordian stage of the Late Jurassic period in southern England, at a time when Europe was a series of scattered islands.

Megalosauridae is a monophyletic family of carnivorous theropod dinosaurs within the group Megalosauroidea. Appearing in the Middle Jurassic, megalosaurids were among the first major radiation of large theropod dinosaurs. They were a relatively primitive group of basal tetanurans containing two main subfamilies, Megalosaurinae and Afrovenatorinae, along with the basal genus Eustreptospondylus, an unresolved taxon which differs from both subfamilies.

Piatnitzkysaurus is a genus of megalosauroid theropod dinosaur that lived approximately 179 to 177 million years ago during the lower part of the Jurassic Period in what is now Argentina. Piatnitzkysaurus was a moderately large, lightly built, bipedal, ground-dwelling carnivore that could grow up to 6.6 m (21.7 ft) long.

Xuanhanosaurus is a genus of theropod dinosaur that lived during the Middle Jurassic (Bathonian) of the Sichuan Basin, China, around 166 million years ago. This taxon represents one of the various non-coelurosaurian tetanuran taxa found on the Middle Jurassic of the region, uncovered in the Lower Shaximiao Formation. Although it has been known for more than 40 years, this taxon has been the subject of very few studies, although most seem to agree that it is a tetanuran, possibly a basal allosauroid, highlighting the fact that it has a vestigial fourth metacarpal.
Chuandongocoelurus is a genus of carnivorous tetanuran theropod dinosaur from the Jurassic of China.

Condorraptor is an extinct genus of megalosauroid theropod dinosaur. Its genus name means 'robber from Cerro Condor', referencing a nearby village, while its species name, currumili, is named after Hipolito Currumil, the landowner and discoverer of the locality. It was among the earliest large South American theropods, having been found in Lower Jurassic strata of the Cañadón Asfalto Formation in the Cañadón Asfalto Basin of Argentina. The type species, described in 2005, is Condorraptor currumili. It is based on a tibia, with an associated partial skeleton that may belong to the same individual. Initially described as a basal tetanuran, Benson (2010) found it to be a piatnitzkysaurid megalosauroid and the sister taxon of Piatnitzkysaurus, a finding supported by later studies.

Metriacanthosauridae is an extinct family of allosauroid theropod dinosaurs that lived from the Middle Jurassic to the Early Cretaceous. When broken down into its Greek roots, it means "moderately-spined lizards". The family is split into two subgroups: Metriacanthosaurinae, which includes dinosaurs closely related to Metriacanthosaurus, and another group composed of the close relatives of Yangchuanosaurus. Metriacanthosaurids are considered carnosaurs, belonging to the Allosauroidea superfamily. The group includes species of large range in body size. Of their physical traits, most notable are their neural spines. The records of the group are mostly confined to Asia, though Metriacanthosaurus is known from Europe. Metriacanthosauridae is used as a senior synonym of Sinraptoridae.

Dubreuillosaurus is a genus of carnivorous dinosaur from the middle Jurassic Period. It is a megalosaurid theropod. Its fossils were found in France. The only named species, Dubreuillosaurus valesdunensis, was originally described as a species of Poekilopleuron, Poekilopleuron? valesdunensis, which is still formally the type species of the genus. It was later renamed Dubreuillosaurus valesdunensis when, in 2005, Allain came to the conclusion that it was not part of the genus Poekilopleuron. Its type specimen, MNHN 1998-13, is only rivalled in the number of preserved elements in this group by that of Eustreptospondylus. Dubreuillosaurus is considered to be the sister species of Magnosaurus. It did not show signs of insular dwarfism even though it was uncovered on an island.

Kaijiangosaurus is a genus of carnivorous tetanuran theropod dinosaur from the Middle Jurassic of China. In 1984 He Xinlu named and described the type species Kaijiangosaurus lini. The generic name refers to the River (jiang) Kai. The specific name honours the paleontologist Lin Wenqiu.

Duriavenator is a genus of theropod dinosaur that lived in what is now England during the Middle Jurassic, about 168 million years ago. In 1882, upper and lower jaw bones of a dinosaur were collected near Sherborne in Dorset, and Richard Owen considered the fossils to belong to the species Megalosaurus bucklandii, the first named non-bird dinosaur. By 1964, the specimen was recognised as belonging to a different species, and in 1974 it was described as a new species of Megalosaurus, M. hesperis; the specific name means 'the West' or 'western'. Later researchers questioned whether the species belonged to Megalosaurus, in which many fragmentary theropods from around the world had historically been placed. After examining the taxonomic issues surrounding Megalosaurus, Roger B. J. Benson moved M. hesperis to its own genus in 2008, Duriavenator; this name means "Dorset hunter".

Megaraptora is a clade of carnivorous theropod dinosaurs. Its derived members, the Megaraptoridae are noted for their large hand claws and powerfully-built forelimbs, which are usually reduced in size in other large theropods.

Haplocheirus is an extinct genus of theropod dinosaur from the Middle Jurassic Shishugou Formation of Xinjiang in China. It is generally considered to be an alvarezsauroid, although some researchers have questioned this assignment. The genus contains a single species, H. sollers, which is known from a mostly complete skeleton including the skull.

Zuolong is an extinct genus of tetanuran theropod from the Late Jurassic period of China. The type and only species is Z. salleei. The generic name of Zuolong is in honor of General Zu Zōngtáng with the Chinese word "long" which means dragon. The specific epithet "salleei" is in honor of Hilmar Sallee, who funded the expedition which led to the specimen's discovery.
Hexing is an extinct genus of basal ornithomimosaur dinosaur known from the Early Cretaceous of northeastern China. It contains a single species, Hexing qingyi.

Aorun is a genus of carnivorous theropod dinosaur first discovered in 2006, with its scientific description published in 2013. It is one of the oldest known coelurosaurian dinosaurs and is estimated to have lived ~161.6 million years ago during the Late Jurassic Period. It is the fifth extinct theropod discovered from Wucaiwan, China.

























