
Deinonychus is a genus of dromaeosaurid theropod dinosaur with one described species, Deinonychus antirrhopus. This species, which could grow up to 3.4 meters (11 ft) long, lived during the early Cretaceous Period, about 115–108 million years ago. Fossils have been recovered from the U.S. states of Montana, Utah, Wyoming, and Oklahoma, in rocks of the Cloverly Formation and Antlers Formation, though teeth that may belong to Deinonychus have been found much farther east in Maryland.

Sauroposeidon is a genus of sauropod dinosaur known from several incomplete specimens including a bone bed and fossilized trackways that have been found in the U.S. states of Oklahoma, Wyoming, and Texas.

Acrocanthosaurus is a genus of carcharodontosaurid dinosaur that existed in what is now North America during the Aptian and early Albian stages of the Early Cretaceous, from 113 to 110 million years ago. Like most dinosaur genera, Acrocanthosaurus contains only a single species, A. atokensis. It had a continent-wide range, with fossil remains known from the U.S. states of Oklahoma, Texas, and Wyoming in the west, and Maryland in the east.

John Harold Ostrom was an American paleontologist who revolutionized the modern understanding of dinosaurs. Ostrom's work inspired what his pupil Robert T. Bakker has termed a "dinosaur renaissance".

Sauropelta is a genus of nodosaurid dinosaur that existed in the Early Cretaceous Period of North America. One species has been named although others may have existed. Anatomically, Sauropelta is one of the most well-understood nodosaurids, with fossilized remains recovered in the U.S. states of Wyoming, Montana, and possibly Utah. It is also the earliest known genus of nodosaurinae; most of its remains are found in a section of the Cloverly Formation dated to 108.5 million years ago.
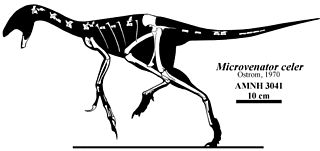
Microvenator is a genus of dinosaur from the Early Cretaceous Cloverly Formation in what is now south central Montana. Microvenator was an oviraptorosaurian theropod. The holotype fossil is an incomplete skeleton, most likely a juvenile with a length of 1.3 m (4.3 ft), and consequently, the adult size remains uncertain. Microvenator celer is primitive and may be the "sister taxon to all other oviraptorosaurs."

Astrodon is a genus of large herbivorous sauropod dinosaur, measuring 20 m (66 ft) in length, 9 m (30 ft) in height and 20 metric tons in body mass. It lived in what is now the eastern United States during the Early Cretaceous period, and fossils have been found in the Arundel Formation, which has been dated through palynomorphs to the Albian about 112 to 110 million years ago.

Albertosaurines, or dinosaurs of the subfamily Albertosaurinae, lived in the Late Cretaceous of United States and Canada. The subfamily was first used by Philip J. Currie, Jørn H. Hurum, and Karol Sabath as a group of tyrannosaurid dinosaurs. It was originally defined as "(Albertosaurus + Gorgosaurus)", including only the two genera. The group is the sister clade to Tyrannosaurinae. In 2007, it was found that the group also contained Maleevosaurus, often synonymized with Tarbosaurus. However, this classification has not been accepted and Maleevosaurus is still considered a juvenile Tarbosaurus or Tyrannosaurus.
The Cedar Mountain Formation is the name given to a distinctive sedimentary geologic formation in eastern Utah, spanning most of the early and mid-Cretaceous. The formation was named for Cedar Mountain in northern Emery County, Utah, where William Lee Stokes first studied the exposures in 1944.

The Cloverly Formation is a geological formation of Early and Late Cretaceous age that is present in parts of Montana, Wyoming, Colorado and Utah in the western United States. It was named for a post office on the eastern side of the Bighorn Basin in Wyoming by N.H. Darton in 1904. The sedimentary rocks of formation were deposited in floodplain environments and contain vertebrate fossils, including a diverse assemblage of dinosaur remains. In 1973, the Cloverly Formation Site was designated as a National Natural Landmark by the National Park Service.
The Antlers Formation is a stratum which ranges from Arkansas through southern Oklahoma into northeastern Texas. The stratum is 150 m (490 ft) thick consisting of silty to sandy mudstone and fine to coarse grained sandstone that is poorly to moderately sorted. The stratum is cemented with clay and calcium carbonate. In places the sandstone may be conglomeratic or ferruginous.

The Eumeralla Formation is a geological formation in Victoria, Australia whose strata date back to the Early Cretaceous. It is Aptian to Albian in age. Dinosaur remains are among the fossils that have been recovered from the formation, particularly from the Dinosaur Cove locality.

Paleontology in Oklahoma refers to paleontological research occurring within or conducted by people from the U.S. state of Oklahoma. Oklahoma has a rich fossil record spanning all three eras of the Phanerozoic Eon. Oklahoma is the best source of Pennsylvanian fossils in the United States due to having an exceptionally complete geologic record of the epoch. From the Cambrian to the Devonian, all of Oklahoma was covered by a sea that would come to be home to creatures like brachiopods, bryozoans, graptolites and trilobites. During the Carboniferous, an expanse of coastal deltaic swamps formed in areas of the state where early tetrapods would leave behind footprints that would later fossilize. The sea withdrew altogether during the Permian period. Oklahoma was home a variety of insects as well as early amphibians and reptiles. Oklahoma stayed dry for most of the Mesozoic. During the Late Triassic, carnivorous dinosaurs left behind footprints that would later fossilize. During the Cretaceous, however, the state was mostly covered by the Western Interior Seaway, which was home to huge ammonites and other marine invertebrates. During the Cenozoic, Oklahoma became home to creatures like bison, camels, creodonts, and horses. During the Ice Age, the state was home to mammoths and mastodons. Local Native Americans are known to have used fossils for medicinal purposes. The Jurassic dinosaur Saurophaganax maximus is the Oklahoma state fossil.

The Trinity Group is a group in the Lower Cretaceous lithostratigraphy of Texas, Arkansas, Mississippi, Louisiana and Oklahoma. It is named for the Trinity River of Texas.

This timeline of dromaeosaurid research is a chronological listing of events in the history of paleontology focused on the dromaeosaurids, a group of sickle-clawed, bird-like theropod dinosaurs including animals like Velociraptor. Since the Native Americans of Montana used the sediments of the Cloverly Formation to produce pigments, they may have encountered remains of the dromaeosaurid Deinonychus hundreds of years before these fossils came to the attention of formally trained scientists.
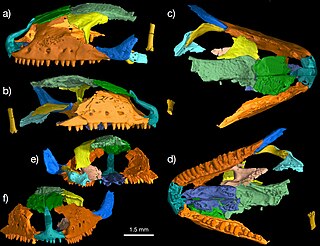
Paramacellodidae is an extinct family of lizards that first appeared in the Middle Jurassic around 170 million years ago (Ma) and became extinct at the end of the Cretaceous around 66 Ma. It was one of the earliest groups of lizards to have undergone an evolutionary radiation, with members found across the supercontinent Laurasia. The phylogenetic relationships and constituent species of Paramacellodidae are uncertain. Many studies regard them to be scincomorphs, a large group that includes skinks and their closest extinct relatives, and possibly also to Cordyoidea, a group that includes spinytail lizards and relatives. Like modern skinks, paramacelloidids had rectangular bony plates called osteoderms covering most of their bodies, including their backs, undersides, and tails. They also had short and robust limbs. Paramacellodids are distinguished from other lizards by the combination two traits in their dentition, the teeth are labiolingually expanded at their bases, and the tooth apices are lingually concave.
Atokasaurus is an extinct genus of scincomorph lizard from the Early Cretaceous of Oklahoma. The type and only species is Atokasaurus metarsiodon, named in 2002 on the basis of a single isolated lower jaw bone found within the Antlers Formation in Atoka County. It is similar in appearance to extinct lizards in the family Paramacellodidae and may itself be a paramacellodid, although the phylogenetic relationships of the group are uncertain. Atokasaurus differs from other paramacellodids in having teeth in the lower jaw with enlarged bases and an S-shaped profile when viewed edge-on.
Ptilotodon is an extinct genus of teiid lizard from the Early Cretaceous of Oklahoma. The type and only known species is Ptilotodon wilsoni, named in 2002 on the basis of a single lower jaw with four teeth found in the Antlers Formation. The small size of the specimen may be an indication that it belonged to a juvenile.















