
Evolution is change in the heritable characteristics of biological populations over successive generations. These characteristics are the expressions of genes that are passed on from parent to offspring during reproduction. Different characteristics tend to exist within any given population as a result of mutation, genetic recombination and other sources of genetic variation. Evolution occurs when evolutionary processes such as natural selection and genetic drift act on this variation, resulting in certain characteristics becoming more common or rare within a population. The evolutionary pressures that determine whether a characteristic should be common or rare within a population constantly change, resulting in the change in heritable characteristics arising over successive generations. It is this process of evolution that has given rise to biodiversity at every level of biological organisation, including the levels of species, individual organisms and molecules.

Ecology is the study of the relationships between living organisms, including humans, and their physical environment. Ecology considers organisms at the individual, population, community, ecosystem, and biosphere level. Ecology overlaps with the closely related sciences of biogeography, evolutionary biology, genetics, ethology, and natural history. Ecology is a branch of biology, and it is not synonymous with environmentalism.
Macroevolution in the modern sense is evolution that is guided by selection among interspecific variation, as opposed to selection among intraspecific variation in microevolution. This modern definition differs from the original concept, which referred macroevolution to the evolution of taxa above the species level.

Cope's rule, named after American paleontologist Edward Drinker Cope, postulates that population lineages tend to increase in body size over evolutionary time. It was never actually stated by Cope, although he favoured the occurrence of linear evolutionary trends. It is sometimes also known as the Cope–Depéret rule, because Charles Depéret explicitly advocated the idea. Theodor Eimer had also done so earlier. The term "Cope's rule" was apparently coined by Bernhard Rensch, based on the fact that Depéret had "lionized Cope" in his book. While the rule has been demonstrated in many instances, it does not hold true at all taxonomic levels, or in all clades. Larger body size is associated with increased fitness for a number of reasons, although there are also some disadvantages both on an individual and on a clade level: clades comprising larger individuals are more prone to extinction, which may act to limit the maximum size of organisms.

Orthogenesis, also known as orthogenetic evolution, progressive evolution, evolutionary progress, or progressionism, is an obsolete biological hypothesis that organisms have an innate tendency to evolve in a definite direction towards some goal (teleology) due to some internal mechanism or "driving force". According to the theory, the largest-scale trends in evolution have an absolute goal such as increasing biological complexity. Prominent historical figures who have championed some form of evolutionary progress include Jean-Baptiste Lamarck, Pierre Teilhard de Chardin, and Henri Bergson.
Evolutionary ecology lies at the intersection of ecology and evolutionary biology. It approaches the study of ecology in a way that explicitly considers the evolutionary histories of species and the interactions between them. Conversely, it can be seen as an approach to the study of evolution that incorporates an understanding of the interactions between the species under consideration. The main subfields of evolutionary ecology are life history evolution, sociobiology, the evolution of interspecific interactions and the evolution of biodiversity and of ecological communities.
A complex adaptive system is a system that is complex in that it is a dynamic network of interactions, but the behavior of the ensemble may not be predictable according to the behavior of the components. It is adaptive in that the individual and collective behavior mutate and self-organize corresponding to the change-initiating micro-event or collection of events. It is a "complex macroscopic collection" of relatively "similar and partially connected micro-structures" formed in order to adapt to the changing environment and increase their survivability as a macro-structure. The Complex Adaptive Systems approach builds on replicator dynamics.
In population genetics and population ecology, population size is the number of individual organisms in a population. Population size is directly associated with amount of genetic drift, and is the underlying cause of effects like population bottlenecks and the founder effect. Genetic drift is the major source of decrease of genetic diversity within populations which drives fixation and can potentially lead to speciation events.
With gene defined as "not just one single physical bit of DNA [but] all replicas of a particular bit of DNA distributed throughout the world", the gene-centered view of evolution, gene's eye view, gene selection theory, or selfish gene theory holds that adaptive evolution occurs through the differential survival of competing genes, increasing the allele frequency of those alleles whose phenotypic trait effects successfully promote their own propagation. The proponents of this viewpoint argue that, since heritable information is passed from generation to generation almost exclusively by DNA, natural selection and evolution are best considered from the perspective of genes.
Devolution, de-evolution, or backward evolution is the notion that species can revert to supposedly more primitive forms over time. The concept relates to the idea that evolution has a purpose (teleology) and is progressive (orthogenesis), for example that feet might be better than hooves or lungs than gills. However, evolutionary biology makes no such assumptions, and natural selection shapes adaptations with no foreknowledge of any kind. It is possible for small changes to be reversed by chance or selection, but this is no different from the normal course of evolution and as such de-evolution is not compatible with a proper understanding of evolution due to natural selection.

Functional ecology is a branch of ecology that focuses on the roles, or functions, that species play in the community or ecosystem in which they occur. In this approach, physiological, anatomical, and life history characteristics of the species are emphasized. The term "function" is used to emphasize certain physiological processes rather than discrete properties, describe an organism's role in a trophic system, or illustrate the effects of natural selective processes on an organism. This sub-discipline of ecology represents the crossroads between ecological patterns and the processes and mechanisms that underlie them. It focuses on traits represented in large number of species and can be measured in two ways – the first being screening, which involves measuring a trait across a number of species, and the second being empiricism, which provides quantitative relationships for the traits measured in screening. Functional ecology often emphasizes an integrative approach, using organism traits and activities to understand community dynamics and ecosystem processes, particularly in response to the rapid global changes occurring in earth's environment.

Full House: The Spread of Excellence from Plato to Darwin is a 1996 book by evolutionary biologist Stephen Jay Gould. It was released in the United Kingdom as Life's Grandeur, with the same subtitle and with an additional eight-page introduction entitled "A Baseball Primer for British Readers".

Andrey Vitalievich Korotayev is a Russian anthropologist, economic historian, comparative political scientist, demographer and sociologist, with major contributions to world-systems theory, cross-cultural studies, Near Eastern history, Big History, and mathematical modelling of social and economic macrodynamics.
The evolution of biological complexity is one important outcome of the process of evolution. Evolution has produced some remarkably complex organisms – although the actual level of complexity is very hard to define or measure accurately in biology, with properties such as gene content, the number of cell types or morphology all proposed as possible metrics.

Dawkins vs. Gould: Survival of the Fittest is a book about the differing views of biologists Richard Dawkins and Stephen Jay Gould by philosopher of biology Kim Sterelny. When first published in 2001 it became an international best-seller. A new edition was published in 2007 to include Gould's The Structure of Evolutionary Theory finished shortly before his death in 2002, and more recent works by Dawkins. The synopsis below is from the 2007 publication.
The Cambrian explosion, Cambrian radiation,Cambrian diversification, or the Biological Big Bang refers to an interval of time approximately 538.8 million years ago in the Cambrian Period when practically all major animal phyla started appearing in the fossil record. It lasted for about 13 – 25 million years and resulted in the divergence of most modern metazoan phyla. The event was accompanied by major diversification in other groups of organisms as well.
Microorganisms engage in a wide variety of social interactions, including cooperation. A cooperative behavior is one that benefits an individual other than the one performing the behavior. This article outlines the various forms of cooperative interactions seen in microbial systems, as well as the benefits that might have driven the evolution of these complex behaviors.

Artificial life is a field of study wherein researchers examine systems related to natural life, its processes, and its evolution, through the use of simulations with computer models, robotics, and biochemistry. The discipline was named by Christopher Langton, an American theoretical biologist, in 1986. In 1987 Langton organized the first conference on the field, in Los Alamos, New Mexico. There are three main kinds of alife, named for their approaches: soft, from software; hard, from hardware; and wet, from biochemistry. Artificial life researchers study traditional biology by trying to recreate aspects of biological phenomena.

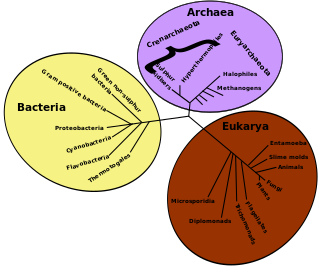
The following outline is provided as an overview of and topical guide to evolution:
The sloshing bucket model of evolution is a theory in evolutionary biology that describes how environmental disturbances varying in magnitude will affect the species present. The theory emphasizes the causal relationship between environmental factors that impinge and affect genealogical systems, providing an overarching view that determines the relationship between the variety of biological systems.