
An action potential occurs when the membrane potential of a specific cell rapidly rises and falls. This depolarization then causes adjacent locations to similarly depolarize. Action potentials occur in several types of excitable cells, which include animal cells like neurons and muscle cells, as well as some plant cells. Certain endocrine cells such as pancreatic beta cells, and certain cells of the anterior pituitary gland are also excitable cells.
Refractoriness is the fundamental property of any object of autowave nature not responding to stimuli, if the object stays in the specific refractory state. In common sense, refractory period is the characteristic recovery time, a period that is associated with the motion of the image point on the left branch of the isocline .

Sir Alan Lloyd Hodgkin was an English physiologist and biophysicist who shared the 1963 Nobel Prize in Physiology or Medicine with Andrew Huxley and John Eccles.
An inhibitory postsynaptic potential (IPSP) is a kind of synaptic potential that makes a postsynaptic neuron less likely to generate an action potential. The opposite of an inhibitory postsynaptic potential is an excitatory postsynaptic potential (EPSP), which is a synaptic potential that makes a postsynaptic neuron more likely to generate an action potential. IPSPs can take place at all chemical synapses, which use the secretion of neurotransmitters to create cell-to-cell signalling. EPSPs and IPSPs compete with each other at numerous synapses of a neuron. This determines whether an action potential occurring at the presynaptic terminal produces an action potential at the postsynaptic membrane. Some common neurotransmitters involved in IPSPs are GABA and glycine.
Hyperpolarization is a change in a cell's membrane potential that makes it more negative. It is the opposite of a depolarization. It inhibits action potentials by increasing the stimulus required to move the membrane potential to the action potential threshold.
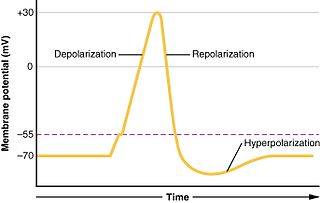
In biology, depolarization or hypopolarization is a change within a cell, during which the cell undergoes a shift in electric charge distribution, resulting in less negative charge inside the cell compared to the outside. Depolarization is essential to the function of many cells, communication between cells, and the overall physiology of an organism.

Membrane potential is the difference in electric potential between the interior and the exterior of a biological cell. It equals the interior potential minus the exterior potential. This is the energy per charge which is required to move a positive charge at constant velocity across the cell membrane from the exterior to the interior.
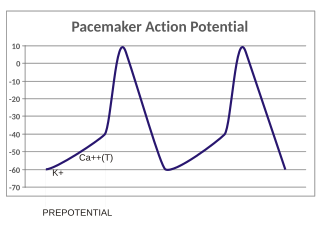
In the pacemaking cells of the heart (e.g., the sinoatrial node), the pacemaker potential (also called the pacemaker current) is the slow, positive increase in voltage across the cell's membrane (the membrane potential) that occurs between the end of one action potential and the beginning of the next action potential. This increase in membrane potential is what causes the cell membrane, which typically maintains a resting membrane potential around -65 mV, to reach the threshold potential and consequently fire the next action potential; thus, the pacemaker potential is what drives the self-generated rhythmic firing (automaticity) of pacemaker cells, and the rate of change (i.e., the slope) of the pacemaker potential is what determines the timing of the next action potential and thus the intrinsic firing rate of the cell. In a healthy sinoatrial node (SAN, a complex tissue within the right atrium containing pacemaker cells that normally determine the intrinsic firing rate for the entire heart), the pacemaker potential is the main determinant of the heart rate. Because the pacemaker potential represents the non-contracting time between heart beats (diastole), it is also called the diastolic depolarization. The amount of net inward current required to move the cell membrane potential during the pacemaker phase is extremely small, in the order of few pAs, but this net flux arises from time to time changing contribution of several currents that flow with different voltage and time dependence. Evidence in support of the active presence of K+, Ca2+, Na+ channels and Na+/K+ exchanger during the pacemaker phase have been variously reported in the literature, but several indications point to the “funny”(If) current as one of the most important.(see funny current). There is now substantial evidence that also sarcoplasmic reticulum (SR) Ca2+-transients participate to the generation of the diastolic depolarization via a process involving the Na–Ca exchanger.

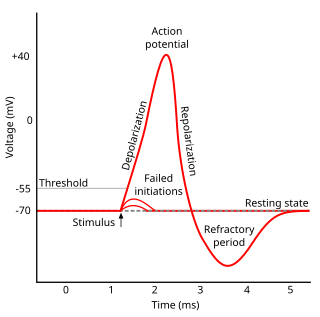
In electrophysiology, the threshold potential is the critical level to which a membrane potential must be depolarized to initiate an action potential. In neuroscience, threshold potentials are necessary to regulate and propagate signaling in both the central nervous system (CNS) and the peripheral nervous system (PNS).

The voltage clamp is an experimental method used by electrophysiologists to measure the ion currents through the membranes of excitable cells, such as neurons, while holding the membrane voltage at a set level. A basic voltage clamp will iteratively measure the membrane potential, and then change the membrane potential (voltage) to a desired value by adding the necessary current. This "clamps" the cell membrane at a desired constant voltage, allowing the voltage clamp to record what currents are delivered. Because the currents applied to the cell must be equal to the current going across the cell membrane at the set voltage, the recorded currents indicate how the cell reacts to changes in membrane potential. Cell membranes of excitable cells contain many different kinds of ion channels, some of which are voltage-gated. The voltage clamp allows the membrane voltage to be manipulated independently of the ionic currents, allowing the current–voltage relationships of membrane channels to be studied.
In physiology, electrotonus refers to the passive spread of charge inside a neuron and between cardiac muscle cells or smooth muscle cells. Passive means that voltage-dependent changes in membrane conductance do not contribute. Neurons and other excitable cells produce two types of electrical potential:
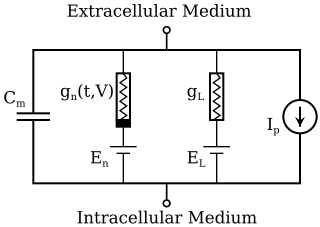
The Hodgkin–Huxley model, or conductance-based model, is a mathematical model that describes how action potentials in neurons are initiated and propagated. It is a set of nonlinear differential equations that approximates the electrical engineering characteristics of excitable cells such as neurons and muscle cells. It is a continuous-time dynamical system.

The soliton hypothesis in neuroscience is a model that claims to explain how action potentials are initiated and conducted along axons based on a thermodynamic theory of nerve pulse propagation. It proposes that the signals travel along the cell's membrane in the form of certain kinds of solitary sound pulses that can be modeled as solitons. The model is proposed as an alternative to the Hodgkin–Huxley model in which action potentials: voltage-gated ion channels in the membrane open and allow sodium ions to enter the cell. The resulting decrease in membrane potential opens nearby voltage-gated sodium channels, thus propagating the action potential. The transmembrane potential is restored by delayed opening of potassium channels. Soliton hypothesis proponents assert that energy is mainly conserved during propagation except dissipation losses; Measured temperature changes are completely inconsistent with the Hodgkin-Huxley model.

Biological neuron models, also known as spiking neuron models, are mathematical descriptions of the conduction of electrical signals in neurons. Neurons are electrically excitable cells within the nervous system, able to fire electric signals, called action potentials, across a neural network. These mathematical models describe the role of the biophysical and geometrical characteristics of neurons on the conduction of electrical activity.
In neurophysiology, several mathematical models of the action potential have been developed, which fall into two basic types. The first type seeks to model the experimental data quantitatively, i.e., to reproduce the measurements of current and voltage exactly. The renowned Hodgkin–Huxley model of the axon from the Loligo squid exemplifies such models. Although qualitatively correct, the H-H model does not describe every type of excitable membrane accurately, since it considers only two ions, each with only one type of voltage-sensitive channel. However, other ions such as calcium may be important and there is a great diversity of channels for all ions. As an example, the cardiac action potential illustrates how differently shaped action potentials can be generated on membranes with voltage-sensitive calcium channels and different types of sodium/potassium channels. The second type of mathematical model is a simplification of the first type; the goal is not to reproduce the experimental data, but to understand qualitatively the role of action potentials in neural circuits. For such a purpose, detailed physiological models may be unnecessarily complicated and may obscure the "forest for the trees". The FitzHugh–Nagumo model is typical of this class, which is often studied for its entrainment behavior. Entrainment is commonly observed in nature, for example in the synchronized lighting of fireflies, which is coordinated by a burst of action potentials; entrainment can also be observed in individual neurons. Both types of models may be used to understand the behavior of small biological neural networks, such as the central pattern generators responsible for some automatic reflex actions. Such networks can generate a complex temporal pattern of action potentials that is used to coordinate muscular contractions, such as those involved in breathing or fast swimming to escape a predator.

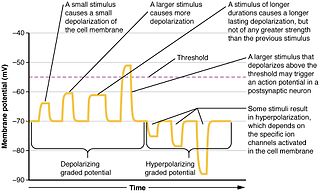
Summation, which includes both spatial summation and temporal summation, is the process that determines whether or not an action potential will be generated by the combined effects of excitatory and inhibitory signals, both from multiple simultaneous inputs, and from repeated inputs. Depending on the sum total of many individual inputs, summation may or may not reach the threshold voltage to trigger an action potential.
Cellular neuroscience is a branch of neuroscience concerned with the study of neurons at a cellular level. This includes morphology and physiological properties of single neurons. Several techniques such as intracellular recording, patch-clamp, and voltage-clamp technique, pharmacology, confocal imaging, molecular biology, two photon laser scanning microscopy and Ca2+ imaging have been used to study activity at the cellular level. Cellular neuroscience examines the various types of neurons, the functions of different neurons, the influence of neurons upon each other, and how neurons work together.
A depolarizing prepulse (DPP) is an electrical stimulus that causes the potential difference measured across a neuronal membrane to become more positive or less negative, and precedes another electrical stimulus. DPPs may be of either the voltage or current stimulus variety and have been used to inhibit neural activity, selectively excite neurons, and increase the pain threshold associated with electrocutaneous stimulation.
Neural accommodation or neuronal accommodation occurs when a neuron or muscle cell is depolarised by slowly rising current in vitro. The Hodgkin–Huxley model also shows accommodation. Sudden depolarisation of a nerve evokes propagated action potential by activating voltage-gated fast sodium channels incorporated in the cell membrane if the depolarisation is strong enough to reach threshold. The open sodium channels allow more sodium ions to flow into the cell and resulting in further depolarisation, which will subsequently open even more sodium channels. At a certain moment this process becomes regenerative and results in the rapid ascending phase of action potential. In parallel with the depolarisation and sodium channel activation, the inactivation process of the sodium channels is also driven by depolarisation. Since the inactivation is much slower than the activation process, during the regenerative phase of action potential, inactivation is unable to prevent the "chain reaction"-like rapid increase in the membrane voltage.
An action potential pulse is a mathematically and experimentally correct Synchronized Oscillating Lipid Pulse coupled with an Action Potential. This is a continuation of Hodgkin Huxley's work in 1952 with the inclusion of accurately modelling ion channel proteins, including their dynamics and speed of activation.