Periarchicortex is one of two subtypes of periallocortex, [1] [2] the other being peripaleocortex. [3] It is formed at borders between archicortex (a subtype of allocortex) and isocortex and shows slow histological transition from the four-layered structure typical for archicortex to the six-layered structure typical for isocortex. [1]
Periallocortex is one of three subtypes of allocortex, the other two subtypes being paleocortex and archicortex. The periallocortex is formed at transition areas where any of the other two subtypes of allocortex borders with the neocortex.
Peripaleocortex is one of two subtypes of periallocortex, the other being periarchicortex. Peripaleocortex is formed at borders between isocortex (neocortex) and paleocortex. It shows slow histological transition from the three-layered structure characteristic of paleocortex to the typical six-layered structure characteristic of isocortex. The main peripaleocortex area is anterior insular cortex.

In anatomy of animals, the archicortex or archipallium is the phylogenetically oldest region of the brain's pallium or cortex.
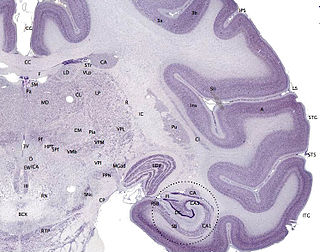
Cortical areas that are generally considered to belong to periarchicortex, include presubiculum, parasubiculum, entorhinal cortex, perirhinal cortex, retrosplenial cortex, periarchicortical part of cingulate cortex, posterior part of subcallosal area, and subgenual area. [1]
In the rodent, the parasubiculum is a retrohippocampal isocortical structure, and a major component of the subicular complex. It receives numerous subcortical and cortical inputs, and sends major projections to the superficial layers of the entorhinal cortex.

The entorhinal cortex (EC) is an area of the brain located in the medial temporal lobe and functioning as a hub in a widespread network for memory, navigation and the perception of time. The EC is the main interface between the hippocampus and neocortex. The EC-hippocampus system plays an important role in declarative (autobiographical/episodic/semantic) memories and in particular spatial memories including memory formation, memory consolidation, and memory optimization in sleep. The EC is also responsible for the pre-processing (familiarity) of the input signals in the reflex nictitating membrane response of classical trace conditioning, the association of impulses from the eye and the ear occurs in the entorhinal cortex.
The Perirhinal cortex is a cortical region in the medial temporal lobe that is made up of Brodmann areas 35 and 36. It receives highly processed sensory information from all sensory regions, and is generally accepted to be an important region for memory. It is bordered caudally by postrhinal cortex or parahippocampal cortex and ventrally and medially by entorhinal cortex.
Periarchicortex does not contact immediately at borders with the true isocortex and does not transit directly into it. Instead, another transitional area from the isocortex side, called proisocortex, is formed at such borders. [4] So, at borders between "true" archicortex and true isocortex, there are two transitional areas. One such area, which is anatomically located closer to the archicortex side and histologically more resembling it, is called periarchicortex. [1] Another transitional area, which is anatomically located closer to the true isocortex and histologically more resembling it, is called proisocortex. [4]
Proisocortex or pro-isocortex is one of two subtypes of cortical areas in the areas belonging to the neocortex. The other subtype is termed the true isocortex. Proisocortical areas are transitional areas placed between areas of true isocortex and areas of periallocortex. The histological structure of proisocortex is also transitional between true isocortex and either peripaleocortex or periarchicortex, depending on with which subtype of periallocortex the given proisocortical area borders.