
Xylem is one of the two types of transport tissue in vascular plants, the other being phloem. The basic function of the xylem is to transport water from roots to stems and leaves, but it also transports nutrients. The word xylem is derived from the Ancient Greek word ξύλον (xylon), meaning "wood"; the best-known xylem tissue is wood, though it is found throughout a plant. The term was introduced by Carl Nägeli in 1858.
Phloem is the living tissue in vascular plants that transports the soluble organic compounds made during photosynthesis and known as photosynthates, in particular the sugar sucrose, to the rest of the plant. This transport process is called translocation. In trees, the phloem is the innermost layer of the bark, hence the name, derived from the Ancient Greek word φλοιός (phloiós), meaning "bark". The term was introduced by Carl Nägeli in 1858. Different types of phloem can be distinguished. The early phloem formed in the growth apices is called protophloem. Protophloem eventually becomes obliterated once it connects to the durable phloem in mature organs, the metaphloem. Further, secondary phloem is formed during the thickening of stem structures.

Vascular plants, also called tracheophytes or collectively tracheophyta, form a large group of land plants that have lignified tissues for conducting water and minerals throughout the plant. They also have a specialized non-lignified tissue to conduct products of photosynthesis. Vascular plants include the clubmosses, horsetails, ferns, gymnosperms, and angiosperms. Scientific names for the group include Tracheophyta, Tracheobionta and Equisetopsida sensu lato. Some early land plants had less developed vascular tissue; the term eutracheophyte has been used for all other vascular plants, including all living ones.

In biology, tissue is an assembly of similar cells and their extracellular matrix from the same embryonic origin that together carry out a specific function. Tissues occupy a biological organizational level between cells and a complete organ. Accordingly, organs are formed by the functional grouping together of multiple tissues.

Bark is the outermost layer of stems and roots of woody plants. Plants with bark include trees, woody vines, and shrubs. Bark refers to all the tissues outside the vascular cambium and is a nontechnical term. It overlays the wood and consists of the inner bark and the outer bark. The inner bark, which in older stems is living tissue, includes the innermost layer of the periderm. The outer bark on older stems includes the dead tissue on the surface of the stems, along with parts of the outermost periderm and all the tissues on the outer side of the periderm. The outer bark on trees which lies external to the living periderm is also called the rhytidome.

The vascular cambium is the main growth tissue in the stems and roots of many plants, specifically in dicots such as buttercups and oak trees, gymnosperms such as pine trees, as well as in certain other vascular plants. It produces secondary xylem inwards, towards the pith, and secondary phloem outwards, towards the bark.
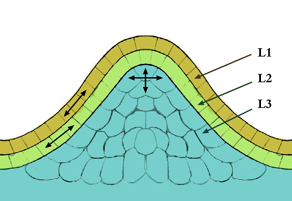
The meristem is a type of tissue found in plants. It consists of undifferentiated cells capable of cell division. Cells in the meristem can develop into all the other tissues and organs that occur in plants. These cells continue to divide until they become differentiated and lose the ability to divide.

In botany, the trunk is the stem and main wooden axis of a tree, which is an important feature in tree identification, and which often differs markedly from the bottom of the trunk to the top, depending on the species.

Lepidodendron is an extinct genus of primitive lycopodian vascular plants belonging the order Lepidodendrales. It is well preserved and common in the fossil record. Like other Lepidodendrales, species of Lepidodendron grew as large-tree-like plants in wetland coal forest environments. They sometimes reached heights of 50 metres, and the trunks were often over 1 m (3.3 ft) in diameter. They are often known as "scale trees", due to their bark having been covered in diamond shaped leaf-bases, from which leaves grew during earlier stages of growth. However, they are correctly defined as arborescent lycophytes. They thrived during the Carboniferous Period, and persisted until the end of the Permian around 252 million years ago. Sometimes erroneously called "giant club mosses", the genus was actually more closely related to modern quillworts than to modern club mosses. In the form classification system used in paleobotany, Lepidodendron is both used for the whole plant as well as specifically the stems and leaves.

Calamites is a genus of extinct arborescent (tree-like) horsetails to which the modern horsetails are closely related. Unlike their herbaceous modern cousins, these plants were medium-sized trees, growing to heights of 30–50 meters. They were components of the understories of coal swamps of the Carboniferous Period.
The pericycle is a cylinder of parenchyma or sclerenchyma cells that lies just inside the endodermis and is the outer most part of the stele of plants.
A vascular bundle is a part of the transport system in vascular plants. The transport itself happens in the stem, which exists in two forms: xylem and phloem. Both these tissues are present in a vascular bundle, which in addition will include supporting and protective tissues. In addition, there is also a tissue between xylem and phloem which is the cambium.

Vascular tissue is a complex conducting tissue, formed of more than one cell type, found in vascular plants. The primary components of vascular tissue are the xylem and phloem. These two tissues transport fluid and nutrients internally. There are also two meristems associated with vascular tissue: the vascular cambium and the cork cambium. All the vascular tissues within a particular plant together constitute the vascular tissue system of that plant.
The ascent of sap in the xylem tissue of plants is the upward movement of water and minerals from the root to the aerial parts of the plant. The conducting cells in xylem are typically non-living and include, in various groups of plants, vessel members and tracheids. Both of these cell types have thick, lignified secondary cell walls and are dead at maturity. Although several mechanisms have been proposed to explain how sap moves through the xylem, the cohesion-tension mechanism has the most support. Although cohesion-tension has received criticism due to the apparent existence of large negative pressures in some living plants, experimental and observational data favor this mechanism.

In botany, secondary growth is the growth that results from cell division in the cambia or lateral meristems and that causes the stems and roots to thicken, while primary growth is growth that occurs as a result of cell division at the tips of stems and roots, causing them to elongate, and gives rise to primary tissue. Secondary growth occurs in most seed plants, but monocots usually lack secondary growth. If they do have secondary growth, it differs from the typical pattern of other seed plants.

Lepidodendrales or arborescent lycophytes are an extinct order of primitive, vascular, heterosporous, arborescent (tree-like) plants belonging to Lycopodiopsida. Members of Lepidodendrales are the best understood of the fossil lycopsids due to the vast diversity of Lepidodendrales specimens and the diversity in which they were preserved; the extensive distribution of Lepidodendrales specimens as well as their well-preservedness lends paleobotanists exceptionally detailed knowledge of the coal-swamp giants’ reproductive biology, vegetative development, and role in their paleoecosystem. The defining characteristics of the Lepidodendrales are their secondary xylem, extensive periderm development, three-zoned cortex, rootlike appendages known as stigmarian rootlets arranged in a spiralling pattern, and megasporangium each containing a single functional megaspore that germinates inside the sporangium. Many of these different plant organs have been assigned both generic and specific names as relatively few have been found organically attached to each other. Some specimens have been discovered which indicate heights of 40 and even 50 meters and diameters of over 2 meters at the base. The massive trunks of some species branched profusely, producing large crowns of leafy twigs; though some leaves were up to 1 meter long, most were much shorter, and when leaves dropped from branches their conspicuous leaf bases remained on the surface of branches. Strobili could be found at the tips of distal branches or in an area at the top of the main trunk. The underground organs of Lepidodendrales typically consisted of dichotomizing axes bearing helically arranged, lateral appendages serving an equivalent function to roots. Sometimes called "giant club mosses", they are believed to be more closely related to extant quillworts based on xylem, although fossil specimens of extinct Selaginellales from the Late Carboniferous also had secondary xylem.

A woody plant is a plant that produces wood as its structural tissue and thus has a hard stem. In cold climates, woody plants further survive winter or dry season above ground, as opposed to herbaceous plants that die back to the ground until spring.

A stem is one of two main structural axes of a vascular plant, the other being the root. It supports leaves, flowers and fruits, transports water and dissolved substances between the roots and the shoots in the xylem and phloem, photosynthesis takes place here, stores nutrients, and produces new living tissue. The stem can also be called halm or haulm or culms.
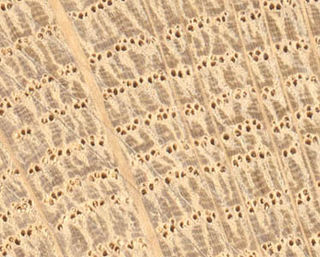
Medullary rays, also known as vascular rays or pith rays, are cellular structures found in some species of wood. They appear as radial planar structures, perpendicular to the growth rings, which are visible to the naked eye. In a transverse section they appear as radiating lines from the centre of the log. In an axial section they may appear as a variety of transverse markings, depending on how close the section is to the plane of the ray. In a tangential section they may be hard to see at all.

A cambium, in plants, is a tissue layer that provides partially undifferentiated cells for plant growth. It is found in the area between xylem and phloem. A cambium can also be defined as a cellular plant tissue from which phloem, xylem, or cork grows by division, resulting in secondary thickening. It forms parallel rows of cells, which result in secondary tissues.