
The serotonin transporter also known as the sodium-dependent serotonin transporter and solute carrier family 6 member 4 is a protein that in humans is encoded by the SLC6A4 gene. SERT is a type of monoamine transporter protein that transports the neurotransmitter serotonin from the synaptic cleft back to the presynaptic neuron, in a process known as serotonin reuptake.
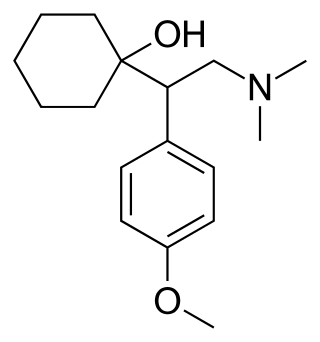
Venlafaxine, sold under the brand name Effexor among others, is an antidepressant medication of the serotonin–norepinephrine reuptake inhibitor (SNRI) class. It is used to treat major depressive disorder, generalized anxiety disorder, panic disorder, and social anxiety disorder. Studies have shown that venlafaxine improves post-traumatic stress disorder (PTSD). It may also be used for chronic pain. It is taken orally. It is also available as the salt venlafaxine besylate in an extended-release formulation.

Catechol-O-methyltransferase is one of several enzymes that degrade catecholamines, catecholestrogens, and various drugs and substances having a catechol structure. In humans, catechol-O-methyltransferase protein is encoded by the COMT gene. Two isoforms of COMT are produced: the soluble short form (S-COMT) and the membrane bound long form (MB-COMT). As the regulation of catecholamines is impaired in a number of medical conditions, several pharmaceutical drugs target COMT to alter its activity and therefore the availability of catecholamines. COMT was first discovered by the biochemist Julius Axelrod in 1957.
Generalized anxiety disorder (GAD) is a mental and behavioral disorder, specifically an anxiety disorder characterized by excessive, uncontrollable and often irrational worry about events or activities. Worry often interferes with daily functioning, and individuals with GAD are often overly concerned about everyday matters such as health, finances, death, family, relationship concerns, or work difficulties. Symptoms may include excessive worry, restlessness, trouble sleeping, exhaustion, irritability, sweating, and trembling.
The candidate gene approach to conducting genetic association studies focuses on associations between genetic variation within pre-specified genes of interest, and phenotypes or disease states. This is in contrast to genome-wide association studies (GWAS), which is a hypothesis-free approach that scans the entire genome for associations between common genetic variants and traits of interest. Candidate genes are most often selected for study based on a priori knowledge of the gene's biological functional impact on the trait or disease in question. The rationale behind focusing on allelic variation in specific, biologically relevant regions of the genome is that certain alleles within a gene may directly impact the function of the gene in question and lead to variation in the phenotype or disease state being investigated. This approach often uses the case-control study design to try to answer the question, "Is one allele of a candidate gene more frequently seen in subjects with the disease than in subjects without the disease?" Candidate genes hypothesized to be associated with complex traits have generally not been replicated by subsequent GWASs or highly powered replication attempts. The failure of candidate gene studies to shed light on the specific genes underlying such traits has been ascribed to insufficient statistical power, low prior probability that scientists can correctly guess a specific allele within a specific gene that is related to a trait, poor methodological practices, and data dredging.

Beck's cognitive triad, also known as the negative triad, is a cognitive-therapeutic view of the three key elements of a person's belief system present in depression. It was proposed by Aaron Beck in 1967. The triad forms part of his cognitive theory of depression and the concept is used as part of CBT, particularly in Beck's "Treatment of Negative Automatic Thoughts" (TNAT) approach.
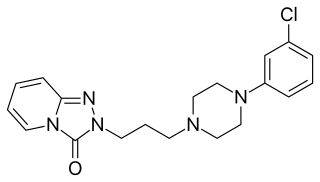
Trazodone, sold under many brand names, is an antidepressant medication. It is used to treat major depressive disorder, anxiety disorders, and insomnia. The medication is taken orally.

Gene–environment interaction is when two different genotypes respond to environmental variation in different ways. A norm of reaction is a graph that shows the relationship between genes and environmental factors when phenotypic differences are continuous. They can help illustrate GxE interactions. When the norm of reaction is not parallel, as shown in the figure below, there is a gene by environment interaction. This indicates that each genotype responds to environmental variation in a different way. Environmental variation can be physical, chemical, biological, behavior patterns or life events.

Monoamine oxidase A, also known as MAO-A, is an enzyme that in humans is encoded by the MAOA gene. This gene is one of two neighboring gene family members that encode mitochondrial enzymes which catalyze the oxidative deamination of amines, such as dopamine, norepinephrine, and serotonin. A mutation of this gene results in Brunner syndrome. This gene has also been associated with a variety of other psychiatric disorders, including antisocial behavior. Alternatively spliced transcript variants encoding multiple isoforms have been observed.
Neuroticism is a personality trait associated with negative emotions. It is one of the Big Five traits. Individuals with high scores on neuroticism are more likely than average to experience such feelings as anxiety, worry, fear, anger, frustration, envy, jealousy, pessimism, guilt, depressed mood, and loneliness. Such people are thought to respond worse to stressors and are more likely to interpret ordinary situations, such as minor frustrations, as appearing hopelessly difficult. Their behavioral responses may include procrastination, substance use, and other maladaptive behaviors, which may aid in relieving negative emotions and generating positive ones.

DASB, also known as 3-amino-4-(2-dimethylaminomethylphenylsulfanyl)-benzonitrile, is a compound that binds to the serotonin transporter. Labeled with carbon-11 — a radioactive isotope — it has been used as a radioligand in neuroimaging with positron emission tomography (PET) since around year 2000. In this context it is regarded as one of the superior radioligands for PET study of the serotonin transporter in the brain, since it has high selectivity for the serotonin transporter.
In genetics, rs6313 also called T102C or C102T is a gene variation—a single nucleotide polymorphism (SNP)—in the human HTR2A gene that codes for the 5-HT2A receptor. The SNP is a synonymous substitution located in exon 1 of the gene where it is involved in coding the 34th amino acid as serine.
In genetics, rs6311 is a gene variation—a single nucleotide polymorphism (SNP)—in the human HTR2A gene that codes for the 5-HT2A receptor. 5-HT2A is a neuroreceptor, and several scientific studies have investigated the effect of the genetic variation on personality, e.g., personality traits measured with the Temperament and Character Inventory or with a psychological task measuring impulsive behavior. The SNP has also been investigated in rheumatology studies.
rs6295, also called C(-1019)G, is a gene variation—a single nucleotide polymorphism (SNP)—in the HTR1A gene. It is one of the most investigated SNPs of its gene. The C-allele is the most prevalent with 0.675 against the G-allele with 0.325 among Caucasian.
Rs6265, also called Val66Met or G196A, is a gene variation, a single nucleotide polymorphism (SNP) in the BDNF gene that codes for brain-derived neurotrophic factor.

Klaus-Peter Lesch is a German clinical psychiatrist who has been investigating the neurobiological foundation of personality traits.
Scientific studies have found that different brain areas show altered activity in humans with major depressive disorder (MDD), and this has encouraged advocates of various theories that seek to identify a biochemical origin of the disease, as opposed to theories that emphasize psychological or situational causes. Factors spanning these causative groups include nutritional deficiencies in magnesium, vitamin D, and tryptophan with situational origin but biological impact. Several theories concerning the biologically based cause of depression have been suggested over the years, including theories revolving around monoamine neurotransmitters, neuroplasticity, neurogenesis, inflammation and the circadian rhythm. Physical illnesses, including hypothyroidism and mitochondrial disease, can also trigger depressive symptoms.
Terrie Edith Moffitt is an American-British clinical psychologist who is best known for her pioneering research on the development of antisocial behavior and for her collaboration with colleague and partner Avshalom Caspi in research on gene-environment interactions in mental disorders.
The genetic influences of post-traumatic stress disorder (PTSD) are not understood well due to the limitations of any genetic study of mental illness; in that, it cannot be ethically induced in selected groups. Because of this, all studies must use naturally occurring groups with genetic similarities and differences, thus the amount of data is limited. Still, genetics play some role in the development of PTSD.

Bipolar disorder is an affective disorder characterized by periods of elevated and depressed mood. The cause and mechanism of bipolar disorder is not yet known, and the study of its biological origins is ongoing. Although no single gene causes the disorder, a number of genes are linked to increase risk of the disorder, and various gene environment interactions may play a role in predisposing individuals to developing bipolar disorder. Neuroimaging and postmortem studies have found abnormalities in a variety of brain regions, and most commonly implicated regions include the ventral prefrontal cortex and amygdala. Dysfunction in emotional circuits located in these regions have been hypothesized as a mechanism for bipolar disorder. A number of lines of evidence suggests abnormalities in neurotransmission, intracellular signalling, and cellular functioning as possibly playing a role in bipolar disorder.

