
Sexual selection is a mode of natural selection in which members of one biological sex choose mates of the other sex to mate with, and compete with members of the same sex for access to members of the opposite sex. These two forms of selection mean that some individuals have greater reproductive success than others within a population, for example because they are more attractive or prefer more attractive partners to produce offspring. Successful males benefit from frequent mating and monopolizing access to one or more fertile females. Females can maximise the return on the energy they invest in reproduction by selecting and mating with the best males.

Heritability is a statistic used in the fields of breeding and genetics that estimates the degree of variation in a phenotypic trait in a population that is due to genetic variation between individuals in that population. The concept of heritability can be expressed in the form of the following question: "What is the proportion of the variation in a given trait within a population that is not explained by the environment or random chance?"

Sexual dimorphism is the condition where sexes of the same species exhibit different morphological characteristics, particularly characteristics not directly involved in reproduction. The condition occurs in most dioecious species, which consist of most animals and some plants. Differences may include secondary sex characteristics, size, weight, color, markings, or behavioral or cognitive traits. Male-male reproductive competition has evolved a diverse array of sexually dimorphic traits. Aggressive utility traits such as "battle" teeth and blunt heads reinforced as battering rams are used as weapons in aggressive interactions between rivals. Passive displays such as ornamental feathering or song-calling have also evolved mainly through sexual selection. These differences may be subtle or exaggerated and may be subjected to sexual selection and natural selection. The opposite of dimorphism is monomorphism, when both biological sexes are phenotypically indistinguishable from each other.
A maternal effect is a situation where the phenotype of an organism is determined not only by the environment it experiences and its genotype, but also by the environment and genotype of its mother. In genetics, maternal effects occur when an organism shows the phenotype expected from the genotype of the mother, irrespective of its own genotype, often due to the mother supplying messenger RNA or proteins to the egg. Maternal effects can also be caused by the maternal environment independent of genotype, sometimes controlling the size, sex, or behaviour of the offspring. These adaptive maternal effects lead to phenotypes of offspring that increase their fitness. Further, it introduces the concept of phenotypic plasticity, an important evolutionary concept. It has been proposed that maternal effects are important for the evolution of adaptive responses to environmental heterogeneity.

In animals, viviparity is development of the embryo inside the body of the mother, with the maternal circulation providing for the metabolic needs of the embryo's development, until the mother gives birth to a fully or partially developed juvenile that is at least metabolically independent. This is opposed to oviparity, where the embryos develop independently outside the mother in eggs until they are developed enough to break out as hatchlings; and ovoviviparity, where the embryos are developed in eggs that remain carried inside the mother's body until the hatchlings emerge from the mother as juveniles, similar to a live birth.

In evolutionary biology, the Baldwin effect describes an effect of learned behaviour on evolution. James Mark Baldwin and others suggested that an organism's ability to learn new behaviours will affect its reproductive success and will therefore have an effect on the genetic makeup of its species through natural selection. It posits that subsequent selection might reinforce the originally learned behaviors, if adaptive, into more in-born, instinctive ones. Though this process appears similar to Lamarckism, that view proposes that living things inherited their parents' acquired characteristics. The Baldwin effect only posits that learning ability, which is genetically based, is another variable in / contributor to environmental adaptation. First proposed during the Eclipse of Darwinism in the late 19th century, this effect has been independently proposed several times, and today it is generally recognized as part of the modern synthesis.

Allometry is the study of the relationship of body size to shape, anatomy, physiology and behaviour, first outlined by Otto Snell in 1892, by D'Arcy Thompson in 1917 in On Growth and Form and by Julian Huxley in 1932.

The common side-blotched lizard is a species of side-blotched lizard in the family Phrynosomatidae. The species is native to dry regions of the western United States and northern Mexico. It is notable for having a unique form of polymorphism wherein each of the three different male morphs utilizes a different strategy in acquiring mates. The three morphs compete against each other following a pattern of rock paper scissors, where one morph has advantages over another but is outcompeted by the third.
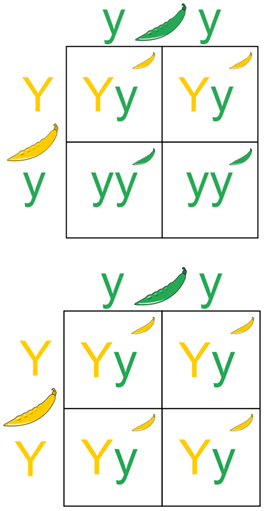
Under the law of dominance in genetics, an individual expressing a dominant phenotype could contain either two copies of the dominant allele or one copy of each dominant and recessive allele. By performing a test cross, one can determine whether the individual is heterozygous or homozygous dominant.

Sexual conflict or sexual antagonism occurs when the two sexes have conflicting optimal fitness strategies concerning reproduction, particularly over the mode and frequency of mating, potentially leading to an evolutionary arms race between males and females. In one example, males may benefit from multiple matings, while multiple matings may harm or endanger females, due to the anatomical differences of that species. Sexual conflict underlies the evolutionary distinction between male and female.

Mate choice is one of the primary mechanisms under which evolution can occur. It is characterized by a "selective response by animals to particular stimuli" which can be observed as behavior. In other words, before an animal engages with a potential mate, they first evaluate various aspects of that mate which are indicative of quality—such as the resources or phenotypes they have—and evaluate whether or not those particular trait(s) are somehow beneficial to them. The evaluation will then incur a response of some sort.

Evolutionary physiology is the study of the biological evolution of physiological structures and processes; that is, the manner in which the functional characteristics of individuals in a population of organisms have responded to natural selection across multiple generations during the history of the population. It is a sub-discipline of both physiology and evolutionary biology. Practitioners in the field come from a variety of backgrounds, including physiology, evolutionary biology, ecology, and genetics.
Rensch's rule is a biological rule on allometrics, concerning the relationship between the extent of sexual size dimorphism and which sex is larger. Across species within a lineage, size dimorphism increases with increasing body size when the male is the larger sex, and decreases with increasing average body size when the female is the larger sex. The rule was proposed by the evolutionary biologist Bernhard Rensch in 1950.

The red-collared widowbird is a species of bird in the family Ploceidae. Red-collared widowbirds are found in grasslands and bush clearings in western and southern Africa. They are known for their long tails and brilliant red badges, both which act as sexual ornaments. They are often associated with other widowbird and bishop species. They are polygynous, where males acquisition of territory is an important determinant in their access to mates. Red-collared widowbirds have a wide range and there is little concern in terms of conservation status.

The long-tailed widowbird is a species of bird in the family Ploceidae. The species are found in Angola, Botswana, the Democratic Republic of the Congo, Kenya, Lesotho, South Africa, Eswatini, and Zambia. The long-tailed widowbird is a medium-sized bird and one of the most common in the territories it inhabits. Adult breeding males are almost entirely black with orange and white shoulders (epaulets), long, wide tails, and a bluish white bill. Females are rather inconspicuous, their feathers streaked tawny and black with pale patches on the chest, breast and back, narrow tail feathers, and horn-colored bills.

Scathophaga stercoraria, commonly known as the yellow dung fly or the golden dung fly, is one of the most familiar and abundant flies in many parts of the Northern Hemisphere. As its common name suggests, it is often found on the feces of large mammals, such as horses, cattle, sheep, deer, and wild boar, where it goes to breed. The distribution of S. stercoraria is likely influenced by human agriculture, especially in northern Europe and North America. The Scathophaga are integral in the animal kingdom due to their role in the natural decomposition of dung in fields. They are also very important in the scientific world due to their short life cycles and susceptibility to experimental manipulations; thus, they have contributed significant knowledge about animal behavior.
A biological ornament is a characteristic of an animal that appears to serve a decorative function rather than a utilitarian function. Many are secondary sexual characteristics, and others appear on young birds during the period when they are dependent on being fed by their parents. Ornaments are used in displays to attract mates, which may lead to the evolutionary process known as sexual selection. An animal may shake, lengthen, or spread out its ornament in order to get the attention of the opposite sex, which will in turn choose the most attractive one with which to mate. Ornaments are most often observed in males, and choosing an extravagantly ornamented male benefits females as the genes that produce the ornament will be passed on to her offspring, increasing their own reproductive fitness. As Ronald Fisher noted, the male offspring will inherit the ornament while the female offspring will inherit the preference for said ornament, which can lead to a positive feedback loop known as a Fisherian runaway. These structures serve as cues to animal sexual behaviour, that is, they are sensory signals that affect mating responses. Therefore, ornamental traits are often selected by mate choice.

Transgenerational epigenetic inheritance is the transmission of epigenetic markers and modifications from one generation to multiple subsequent generations without altering the primary structure of DNA. Thus, the regulation of genes via epigenetic mechanisms can be heritable; the amount of transcripts and proteins produced can be altered by inherited epigenetic changes. In order for epigenetic marks to be heritable, however, they must occur in the gametes in animals, but since plants lack a definitive germline and can propagate, epigenetic marks in any tissue can be heritable.

Phenotypic integration is a metric for measuring the correlation of multiple functionally-related traits to each other. Complex phenotypes often require multiple traits working together in order to function properly. Phenotypic integration is significant because it provides an explanation as to how phenotypes are sustained by relationships between traits. Every organism's phenotype is integrated, organized, and a functional whole. Integration is also associated with functional modules. Modules are complex character units that are tightly associated, such as a flower. It is hypothesized that organisms with high correlations between traits in a module have the most efficient functions. The fitness of a particular value for one phenotypic trait frequently depends on the value of the other phenotypic traits, making it important for those traits evolve together. One trait can have a direct effect on fitness, and it has been shown that the correlations among traits can also change fitness, causing these correlations to be adaptive, rather than solely genetic. Integration can be involved in multiple aspects of life, not just at the genetic level, but during development, or simply at a functional level.
Countergradient variation is a type of phenotypic plasticity that occurs when the phenotypic variation determined by a biological population's genetic components opposes the phenotypic variation caused by an environmental gradient. This can cause different populations of the same organism to display similar phenotypes regardless of their underlying genetics and differences in their environments.

