Bonnie Lynn Bassler is an American molecular biologist; the Squibb Professor in Molecular Biology and chair of the Department of Molecular Biology at Princeton University; and a Howard Hughes Medical Institute Investigator. She has researched cell-to-cell chemical communication in bacteria and discovered key insights into the mechanism by which bacteria communicate, known as quorum sensing. She has contributed to the idea that disruption of chemical signaling can be used as an antimicrobial therapy.
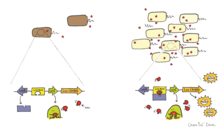
In biology, quorum sensing or quorum signaling (QS) is the ability to detect and respond to cell population density by gene regulation. Quorum sensing is a type of cellular signaling, and more specifically can be considered a type of paracrine signaling. However, it also contains traits of both autocrine signaling: a cell produces both the autoinducer molecule and the receptor for the autoinducer. As one example, QS enables bacteria to restrict the expression of specific genes to the high cell densities at which the resulting phenotypes will be most beneficial, especially for phenotypes that would be ineffective at low cell densities and therefore too energetically costly to express. Many species of bacteria use quorum sensing to coordinate gene expression according to the density of their local population. In a similar fashion, some social insects use quorum sensing to determine where to nest. Quorum sensing in pathogenic bacteria activates host immune signaling and prolongs host survival, by limiting the bacterial intake of nutrients, such as tryptophan, which further is converted to serotonin. As such, quorum sensing allows a commensal interaction between host and pathogenic bacteria. Quorum sensing may also be useful for cancer cell communications.

Bioluminescence is the production and emission of light by living organisms. It is a form of chemiluminescence. Bioluminescence occurs widely in marine vertebrates and invertebrates, as well as in some fungi, microorganisms including some bioluminescent bacteria, and terrestrial arthropods such as fireflies. In some animals, the light is bacteriogenic, produced by symbiotic bacteria such as those from the genus Vibrio; in others, it is autogenic, produced by the animals themselves.

Luciferase is a generic term for the class of oxidative enzymes that produce bioluminescence, and is usually distinguished from a photoprotein. The name was first used by Raphaël Dubois who invented the words luciferin and luciferase, for the substrate and enzyme, respectively. Both words are derived from the Latin word lucifer, meaning "lightbearer", which in turn is derived from the Latin words for "light" (lux) and "to bring or carry" (ferre).

Aliivibrio fischeri is a Gram-negative, rod-shaped bacterium found globally in marine environments. This species has bioluminescent properties, and is found predominantly in symbiosis with various marine animals, such as the Hawaiian bobtail squid. It is heterotrophic, oxidase-positive, and motile by means of a single polar flagella. Free-living A. fischeri cells survive on decaying organic matter. The bacterium is a key research organism for examination of microbial bioluminescence, quorum sensing, and bacterial-animal symbiosis. It is named after Bernhard Fischer, a German microbiologist.

Vibrio harveyi is a Gram-negative, bioluminescent, marine bacterium in the genus Vibrio. V. harveyi is rod-shaped, motile, facultatively anaerobic, halophilic, and competent for both fermentative and respiratory metabolism. It does not grow below 4 °C. V. harveyi can be found free-swimming in tropical marine waters, commensally in the gut microflora of marine animals, and as both a primary and opportunistic pathogen of marine animals, including Gorgonian corals, oysters, prawns, lobsters, the common snook, barramundi, turbot, milkfish, and seahorses. It is responsible for luminous vibriosis, a disease that affects commercially farmed penaeid prawns. Additionally, based on samples taken by ocean-going ships, V. harveyi is thought to be the cause of the milky seas effect, in which, during the night, a uniform blue glow is emitted from the seawater. Some glows can cover nearly 6,000 sq mi (16,000 km2).

N-Acyl homoserine lactones are a class of signaling molecules involved in bacterial quorum sensing, a means of communication between bacteria enabling behaviors based on population density.
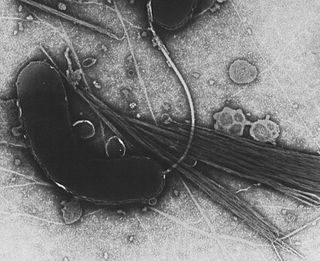
The Vibrionaceae are a family of Pseudomonadota given their own order, Vibrionales. Inhabitants of fresh or salt water, several species are pathogenic, including the type species Vibrio cholerae, which is the agent responsible for cholera. Most bioluminescent bacteria belong to this family, and are typically found as symbionts of deep-sea animals.
Luminescent bacteria emit light as the result of a chemical reaction during which chemical energy is converted to light energy. Luminescent bacteria exist as symbiotic organisms carried within a larger organism, such as many deep sea organisms, including the Lantern Fish, the Angler fish, certain jellyfish, certain clams and the Gulper eel. The light is generated by an enzyme-catalyzed chemoluminescence reaction, wherein the pigment luciferin is oxidised by the enzyme luciferase. The expression of genes related to bioluminescence is controlled by an operon called the lux operon.
Bioluminescence imaging (BLI) is a technology developed over the past decades (1990's and onward). that allows for the noninvasive study of ongoing biological processes Recently, bioluminescence tomography (BLT) has become possible and several systems have become commercially available. In 2011, PerkinElmer acquired one of the most popular lines of optical imaging systems with bioluminescence from Caliper Life Sciences.

Euprymna scolopes, also known as the Hawaiian bobtail squid, is a species of bobtail squid in the family Sepiolidae native to the central Pacific Ocean, where it occurs in shallow coastal waters off the Hawaiian Islands and Midway Island. The type specimen was collected off the Hawaiian Islands and is located at the National Museum of Natural History in Washington, D.C.
Autoinducers are signaling molecules that are produced in response to changes in cell-population density. As the density of quorum sensing bacterial cells increases so does the concentration of the autoinducer. Detection of signal molecules by bacteria acts as stimulation which leads to altered gene expression once the minimal threshold is reached. Quorum sensing is a phenomenon that allows both Gram-negative and Gram-positive bacteria to sense one another and to regulate a wide variety of physiological activities. Such activities include symbiosis, virulence, motility, antibiotic production, and biofilm formation. Autoinducers come in a number of different forms depending on the species, but the effect that they have is similar in many cases. Autoinducers allow bacteria to communicate both within and between different species. This communication alters gene expression and allows bacteria to mount coordinated responses to their environments, in a manner that is comparable to behavior and signaling in higher organisms. Not surprisingly, it has been suggested that quorum sensing may have been an important evolutionary milestone that ultimately gave rise to multicellular life forms.

Autoinducer-2 (AI-2), a furanosyl borate diester or tetrahydroxy furan, is a member of a family of signaling molecules used in quorum sensing. AI-2 is one of only a few known biomolecules incorporating boron. First identified in the marine bacterium Vibrio harveyi, AI-2 is produced and recognized by many Gram-negative and Gram-positive bacteria. AI-2 arises by the reaction of 4,5-dihydroxy-2,3-pentanedione, which is produced enzymatically, with boric acid and is recognized by the two-component sensor kinase LuxPQ in Vibrionaceae.

John Woodland "Woody" Hastings, was a leader in the field of photobiology, especially bioluminescence, and was one of the founders of the field of circadian biology. He was the Paul C. Mangelsdorf Professor of Natural Sciences and Professor of Molecular and Cellular Biology at Harvard University. He published over 400 papers and co-edited three books.
Interspecies quorum sensing is a type of quorum sensing in which bacteria send and receive signals to other species besides their own. This is accomplished by the secretion of signaling molecules which trigger a response in nearby bacteria at high enough concentrations. Once the molecule hits a certain concentration it triggers the transcription of certain genes such as virulence factors. It has been discovered that bacteria can not only interact via quorum sensing with members of their own species but that there is a kind of universal molecule that allows them to gather information about other species as well. This universal molecule is called autoinducer 2 or AI-2.
Vibrio campbellii is a Gram-negative, curved rod-shaped, marine bacterium closely related to its sister species, Vibrio harveyi. It is an emerging pathogen in aquatic organisms.
Everett Peter Greenberg is an American microbiologist. He is the inaugural Eugene and Martha Nester Professor of Microbiology at the Department of Microbiology of the University of Washington School of Medicine. He is best known for his research on quorum sensing, and has received multiple awards for his work.
Margaret McFall-Ngai is an American animal physiologist and biochemist best-known for her work related to the symbiotic relationship between Hawaiian bobtail squid, Euprymna scolopes and bioluminescent bacteria, Vibrio fischeri. Her research helped expand the microbiology field, primarily focused on pathogenicity and decomposition at the time, to include positive microbial associations. She currently is a professor at PBRC’s Kewalo Marine Laboratory and director of the Pacific Biosciences Research Program at the University of Hawaiʻi at Mānoa.
Karen Visick is an American microbiologist and expert in bacterial genetics known for her work on the role of bacteria to form biofilm communities during animal colonization. She conducted doctoral research with geneticist Kelly Hughes at the University of Washington, where she identified a key regulatory checkpoint during construction of the bacterial flagellum. She conducted postdoctoral research on development of the Vibrio fischeri-Euprymna scolopes symbiosis with Ned Ruby at University of Southern California and University of Hawaiʻi. The bacteria are bioluminescent and provide light to the host. Visick and Ruby revealed that bacteria that do not produce light exhibit a defect during host colonization.