
Frontotemporal dementia (FTD), frontotemporal degeneration disease, or frontotemporal neurocognitive disorder encompasses several types of dementia involving the progressive degeneration of the brain's frontal and temporal lobes. FTDs broadly present as behavioral or language disorders with gradual onsets.

Frontotemporal lobar degeneration (FTLD) is a pathological process that occurs in frontotemporal dementia. It is characterized by atrophy in the frontal lobe and temporal lobe of the brain, with sparing of the parietal and occipital lobes.
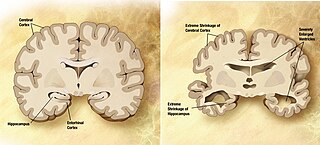
A neurodegenerative disease is caused by the progressive loss of structure or function of neurons, in the process known as neurodegeneration. Such neuronal damage may ultimately involve cell death. Neurodegenerative diseases include amyotrophic lateral sclerosis, multiple sclerosis, Parkinson's disease, Alzheimer's disease, Huntington's disease, multiple system atrophy, tauopathies, and prion diseases. Neurodegeneration can be found in the brain at many different levels of neuronal circuitry, ranging from molecular to systemic. Because there is no known way to reverse the progressive degeneration of neurons, these diseases are considered to be incurable; however research has shown that the two major contributing factors to neurodegeneration are oxidative stress and inflammation. Biomedical research has revealed many similarities between these diseases at the subcellular level, including atypical protein assemblies and induced cell death. These similarities suggest that therapeutic advances against one neurodegenerative disease might ameliorate other diseases as well.

Superoxide dismutase [Cu-Zn] also known as superoxide dismutase 1 or hSod1 is an enzyme that in humans is encoded by the SOD1 gene, located on chromosome 21. SOD1 is one of three human superoxide dismutases. It is implicated in apoptosis, familial amyotrophic lateral sclerosis and Parkinson's disease.

Profilin-1 is a protein that in humans is encoded by the PFN1 gene.

Pur-alpha is a protein that in humans is encoded by the PURA gene located at chromosome 5, band q31.

Alsin is a protein that in humans is encoded by the ALS2 gene. ALS2 orthologs have been identified in all mammals for which complete genome data are available.

TAR DNA-binding protein 43 is a protein that in humans is encoded by the TARDBP gene.

Transmembrane protein 106B is a protein that is encoded by the TMEM106B gene. It is found primarily within neurons and oligodendrocytes in the central nervous system with its subcellular location being in lysosomal membranes. TMEM106B helps facilitate important functions for maintaining a healthy lysosome, and therefore certain mutations and polymorphisms can lead to issues with proper lysosomal function. Lysosomes are in charge of clearing out mis-folded proteins and other debris, and thus, play an important role in neurodegenerative diseases that are driven by the accumulation of various mis-folded proteins and aggregates. Due to its impact on lysosomal function, TMEM106B has been investigated and found to be associated to multiple neurodegenerative diseases.

G2/mitotic-specific cyclin-F is a protein that in humans is encoded by the CCNF gene.

Amyotrophic lateral sclerosis (ALS), also known as motor neurone disease (MND) or Lou Gehrig's disease in the United States, is a rare and terminal neurodegenerative disease that results in the progressive loss of motor neurons that control voluntary muscles. ALS is the most common form of the motor neuron diseases. Early symptoms of ALS include stiff muscles, muscle twitches, gradual increasing weakness, and muscle wasting. Limb-onset ALS begins with weakness in the arms or legs, while bulbar-onset ALS begins with difficulty in speaking or swallowing. Around half of people with ALS develop at least mild difficulties with thinking and behavior, and about 15% develop frontotemporal dementia. Motor neuron loss continues until the abilities to eat, speak, move, or, lastly, breathe are lost.
Project MinE is an independent large scale whole genome research project that was initiated by 2 patients with amyotrophic lateral sclerosis and started on World ALS Day, June 21, 2013.
Multisystem proteinopathy (MSP) is a dominantly inherited, pleiotropic, degenerative disorder of humans that can affect muscle, bone, and/or the central nervous system. MSP can manifest clinically as classical amyotrophic lateral sclerosis (ALS), frontotemporal dementia (FTD), inclusion body myopathy (IBM), Paget's disease of bone (PDB), or as a combination of these disorders. Historically, several different names have been used to describe MSP, most commonly "inclusion body myopathy with early-onset Paget disease and frontotemporal dementia (IBMPFD)" or "inclusion body myopathy with frontotemporal dementia, Paget's disease of bone, and amyotrophic lateral sclerosis (IBMPFD/ALS)." However, IBMPFD and IBMPFD/ALS are now considered outdated classifications and are more properly referred to as MSP, as the disease is clinically heterogeneous and its phenotypic spectrum extends beyond IBM, PDB, FTD, and ALS to include motor neuron disease, Parkinson's disease features, and ataxia features. Although MSP is rare, growing interest in this syndrome derives from the molecular insights the condition provides into the etiological relationship between common age-related degenerative diseases of muscle, bone, and brain.
Coiled-coil-helix-coiled-coil-helix domain-containing protein 10, mitochondrial, also known as Protein N27C7-4 is a protein that in humans is encoded by the CHCHD10 gene.
RNA-dominant diseases are characterized by deleterious mutations that typically result in degenerative disorders affecting various neurological, cardiovascular, and muscular functions. Studies have found that they arise from repetitive non-coding RNA sequences, also known as toxic RNA, which inhibit RNA-binding proteins leading to pathogenic effects. The most studied RNA-dominant diseases include, but are not limited to, myotonic dystrophy and fragile X-associated tremor/ataxia syndrome (FXTAS).

Unc-13 homolog A is a protein that in humans is encoded by the UNC13A gene.
There are more than 25 genes known to be associated with amyotrophic lateral sclerosis (ALS) as of June 2018, which collectively account for about 70% of cases of familial ALS (fALS) and 10% of cases of sporadic ALS (sALS). About 5–10% of cases of ALS are directly inherited. Overall, first-degree relatives of an individual with ALS have a 1% risk of developing ALS. ALS has an oligogenic mode of inheritance, meaning that mutations in two or more genes are required to cause disease.
Research on amyotrophic lateral sclerosis (ALS) has focused on animal models of the disease, its mechanisms, ways to diagnose and track it, and treatments.
Bryan J. Traynor is a neurologist and a senior investigator at the National Institute on Aging, and an adjunct professor at Johns Hopkins University. Dr. Traynor studies the genetics of human neurological conditions such as amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). He led the international consortium that identified pathogenic repeat expansions in the C9orf72 gene as a common cause of ALS and FTD. Dr. Traynor also led efforts that identified other Mendelian genes responsible for familial ALS and dementia, including VCP, MATR3, KIF5A, HTT, and SPTLC1.
Rosa Rademakers is a Dutch neurogeneticist and professor within the Department of Neuroscience at the Mayo Clinic. Her research centers on the genetic basis of neurodegenerative diseases, such as identifying causal genes and their function, exploring familial risk factors, and the mechanism of the degeneration. Her neurodegenerative diseases of focus include "Alzheimer's disease (AD), frontotemporal dementia (FTD) and amyotrophic lateral sclerosis (ALS)." She received a Bachelor of Arts in Biology, a Master of Arts in Biochemistry, and a Ph.D. in Science, all from the University of Antwerp. Originally from the Netherlands, she came to the Mayo Clinic in 2005 for a post-doctoral fellowship, and in 2007 she was given a lab director position.



