Caretaker genes encode products that stabilize the genome. Fundamentally, mutations in caretaker genes lead to genomic instability. Tumor cells arise from two distinct classes of genomic instability: mutational instability arising from changes in the nucleotide sequence of DNA and chromosomal instability arising from improper rearrangement of chromosomes.[1]
Changes in the genome that allow uncontrolled cell proliferation or cell immortality are responsible for cancer. It is believed that the major changes in the genome that lead to cancer arise from mutations in tumor suppressor genes.[2] In 1997, Kinzler and Bert Vogelstein grouped these cancer susceptibility genes into two classes: "caretakers" and "gatekeepers".[3] In 2004, a third classification of tumor suppressor genes was proposed by Franziska Michor, Yoh Iwasa, and Martin Nowak; "landscaper" genes. In contrast to caretaker genes, gatekeeper genes encode gene products that act to prevent growth of potential cancer cells and prevent accumulation of mutations that directly lead to increased cellular proliferation.[4][5]
The third classification of genes, the landscapers, encode products that, when mutated, contribute to the neoplastic growth of cells by fostering a stromal environment conducive to unregulated cell proliferation.[6]
Genes in context
Pathways to cancer via the caretakers
The process of DNA replication inherently places cells at risk of acquiring mutations. Thus, caretaker genes are vitally important to cellular health. Rounds of cell replication allow fixation of mutated genes into the genome.[5] Caretaker genes provide genome stability by preventing the accumulation of these mutations.
Loss of function mutations in caretaker genes allow mutations in other genes to survive that can result in increased conversion of a normal cell to a neoplastic cell, a cell that; (1) divides more often than it should or (2) does not die when conditions warrant cell death.[2] Thus, caretaker genes do not directly regulate cell proliferation. Instead, they prevent other mutations from surviving for example by slowing the cell division process to enable DNA repair to complete, or by initiating apoptosis of the cell.[7] In genetic knock-out and rescue experiments, restoration of a caretaker gene from the mutated form to the wildtype version does not limit tumorigenesis.[8] This is because caretaker genes only indirectly contribute to the pathway to cancer.
Cells deficient in a DNA repair process tend to accumulate unrepaired DNA damages. Cells defective in apoptosis tend to survive even with excessive DNA damage, thus permitting replication of the damaged DNA and consequently carcinogenic mutations. Some key caretaker proteins that contribute to cell survival by acting in DNA repair processes when the level of damage is manageable, become executioners by inducing apoptosis when there is excess DNA damage[9].
Inactivation of caretaker genes is environmentally equivalent to exposing the cell to mutagens incessantly.[10] For example, a mutation in a caretaker gene coding for a DNA repair pathway that leads to the inability to properly repair DNA damage could allow uncontrolled cell growth. This is the result of mutations of other genes that accumulate unchecked as a result of faulty gene products encoded by the caretakers.
In addition to providing genomic stability, caretakers also provide chromosomal stability. Chromosomal instability resulting from dysfunctional caretaker genes is the most common form of genetic instability that leads to cancer in humans.[11] In fact, it has been proposed that these caretaker genes are responsible for many hereditary predispositions to cancers.[12]
In individuals predisposed to cancer via mutations in caretaker genes, a total of three subsequent somatic mutations are required to acquire the cancerous phenotype.[12] Mutations must occur in the remaining normal caretaker allele in addition to both alleles of gatekeeper genes within that cell for the said cell to turn to neoplasia. Thus, the risk of cancer in these affected populations is much less when compared to cancer risk in families predisposed to cancer via the gatekeeper pathway.[12]
Pathways to cancer via the gatekeepers
In many cases, gatekeeper genes encode a system of checks and balances that monitor cell division and death.[4] When tissue damage occurs, for example, products of gatekeeper genes ensure that balance of cell growth over cellular death remains in check.[13] In the presence of competent gatekeeper genes, mutations of other genes do not lead to on-going growth imbalances.
Mutations altering these genes lead to irregular growth regulation and differentiation.[11] Each cell type has only one, or at least only very few, gatekeeper genes.[12] If a person is predisposed to cancer, they have inherited a mutation in one of two copies of a gatekeeper gene. Mutation of the alternate allele leads to progression to neoplasia.[12]
Historically, the term gatekeeper gene was first coined in association with the APC gene, a tumor suppressor that is consistently found to be mutated in colorectal tumors.[14] Gatekeeper genes are in fact specific to the tissues in which they reside.[3]
The probability that mutations occur in other genes increases when DNA repair pathway mechanisms are damaged as a result of mutations in caretaker genes. Thus, the probability that a mutation will take place in a gatekeeper gene increases when the caretaker gene has been mutated.[15]
Apoptosis, or induced cell suicide, usually serves as a mechanism to prevent excessive cellular growth. Gatekeeper genes regulate apoptosis. However, in instances where tissue growth or regrowth is warranted, these signals must be inactivated or net tissue regeneration would be impossible. Thus, mutations in growth-controlling genes would lead to the characteristics of uncontrolled cellular proliferation, neoplasia, while in a parallel cell that had no mutations in the gatekeeper function, simple cell death would ensue.[13]
Pathways to cancer via the landscapers
A third group of genes in which mutations lead to a significant susceptibility to cancer is the class of landscaper genes. Products encoded by landscaper genes do not directly affect cellular growth, but when mutated, contribute to the neoplastic growth of cells by fostering stromal environments conducive to unregulated cell proliferation.[6]
Cells communicate with each other via the ECM through both direct contact and through signaling molecules.[16] Stromal cell abnormalities arising from gene products coded by faulty landscaper genes could induce abnormal cell growth on the epithelium, leading to cancer of that tissue.[17]
Biochemical cascades consisting of signaling proteins occur in the ECM and play an important role to the regulation of many aspects of cell life.[5] Landscaper genes encode products that determine the composition of the membranes in which cells live. For example, large molecular weight glycoproteins and proteoglycans have been found to in association with signaling and structural roles.[5] There exist proteolytic molecules in the ECM that are essential for clearing unwanted molecules, such as growth factors, cell adhesion molecules, and others from the space surrounding cells.[5] It is proposed that landscaper genes control the mechanisms by which these factors are properly cleared. Different characteristics of these membranes lead to different cellular effects, such as differing rates of cell proliferation or differentiation.[16] If, for example, the ECM is disrupted, incoming cells, such as those of the immune system, can overload the area and release chemical signals that induce abnormal cell proliferation.[16] These conditions lead to an environment conducive to tumor growth and the cancerous phenotype.
Gatekeepers, caretakers, and cellular aging
Because mechanisms that control the accumulation of damage through the lifetime of a cell are essential to longevity, it is logical that caretaker and gatekeeper genes play a significant role in cellular aging. Increased activity of caretaker genes postpones aging, increasing lifespan.[18] This is because of the regulatory function associated with caretaker genes in maintaining the stability of the genome. The actions of caretaker genes contribute to increasing lifespan of the cell.
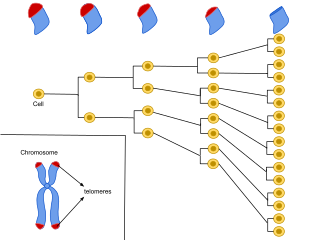
A specific purpose of caretaker genes has been outlined in chromosomal duplication. Caretakers have been identified as crucial to encoding products that maintain the telomeres.[19] It is believed that degradation of telomeres, the ends of chromosomes, through repeated cell cycle divisions, is a main component of cellular aging and death.
It has been suggested that gatekeeper genes confer beneficial anti-cancer affects but may provide deleterious effects that increase aging.[5] This is because young organisms experiencing times of rapid growth necessitate significant anti-cancer mechanisms. As the organism ages, however, these formerly beneficial pathways become deleterious by inducing apoptosis in cells of renewable tissues, causing degeneration of the structure.[5] Studies have shown an increased expression of pro-apoptotic genes in age-related pathologies.[18] This is because the products of gatekeeper genes are directly involved in coding for cellular growth and proliferation.
However, dysfunctional caretaker genes do not always lead to a cancerous phenotype.[18] For example, defects in nucleotide excision repair pathways are associated with premature aging phenotypes in diseases such as Xeroderma pigmentosum and Trichothiodystrophy. These patients exhibit brittle hair, nails, scaly skin, and hearing loss – characteristics associated with simple human aging. This is important because the nucleotide excision repair pathway is a mechanism thought to be encoded by a caretaker gene. Geneticists studying these premature-aging syndromes propose that caretaker genes that determine cell fate also play a significant role in aging.[18] Accumulation of DNA damage with age may be especially prevalent in the central nervous system because of low DNA repair capability in postmitotic brain tissue[20].
Similarly, gatekeeper genes have been identified as having a role in aging disorders that exhibit mutations in such genes without an increased susceptibility to cancer. Experiments with mice that have increased gatekeeper function in the p53 gene show reduced cancer incidence (due to the protective activities of products encoded by p53) but a faster rate of aging.[18]
Cellular senescence, also encoded by a gatekeeper gene, is arrest of the cell cycle in the G1 phase. Qualitative differences have been found between senescent cells and normal cells, including differential expression of cytokines and other factors associated with inflammation.[18] It is believed that this may contribute, in part, to cellular aging.
In sum, although mechanisms encoded by gatekeeper and caretaker genes to protect individuals from cancer early in life, namely induction of apoptosis or senescence, later in life these functions may promote the aging phenotype.[18]
Mutations in context
It has been proposed that mutations in gatekeeper genes could, to an extent, offer a sort of selective advantage to the individual in which the change occurs. This is because cells with these mutations are able to replicate at a faster rate than nearby cells.[4] This is known as "increased somatic fitness".[11] Caretaker genes, on the other hand, confer selective disadvantage because the result is inherently decreased cellular success.[4] However, increased somatic fitness could also arise from a mutation in a caretaker gene if mutations in tumor suppressor genes increase the net reproductive rate of the cell.[11]
Although mutations in gatekeeper genes may lead to the same result as those of caretaker genes, namely cancer, the transcripts that gatekeeper genes encode are significantly different from those encoded by caretaker genes.
In many cases, gatekeeper genes encode a system of checks and balances that monitor cell division and death.[4] In cases of tissue damage, for example, gatekeeper genes would ensure that balance of cell growth over cellular death remains in check.[13] In the presence of competent gatekeeper genes, mutations of other genes would not lead to on-going growth imbalances.
Whether or not mutations in these genes confer beneficial or deleterious effects to the animal depends partially on the environmental context in which these changes occur, a context encoded by the landscaper genes. For example, tissues of the skin and colon reside in compartments of cells that rarely mix with one another.[4] These tissues are replenished by stem cells. Mutations that occur within these cell lineages remain confined to the compartment in which they reside, increasing the future risk of cancer.[4] This is also protective, however, because the cancer will remain confined to that specific area, rather than invading the rest of the body, a phenomenon known as metastasis.
In areas of the body compartmentalized into small subsets of cells, mutations that lead to cancer most often begin with caretaker genes.[4] On the other hand, cancer progression in non-compartmentalized or large cell populations may be a result of initial mutations in gatekeepers.[4]
These delineations offer a suggestion why different types of tissue within the body progress to cancer by differing mechanisms.[4]
Notes
Although the classification of tumor suppressor genes into these categories is helpful to the scientific community, the potential role of many genes cannot be reliably identified as the functions of many genes are rather ill-defined.[3] In some contexts, genes exhibit discrete caretaker function while in other situations gatekeeper characteristics are recognized. An example of one such gene is p53. Patients with Li-Fraumeni syndrome, for example, have mutations in the p53 gene that suggest caretaker function. p53 has an identified role, however, in regulating the cell cycle as well, which is an essential gatekeeper function.[3]
Sources
↑ Van Gent, D. C.; Hoeijmakers, J. H.; Kanaar, R (2001). "Chromosomal stability and the DNA double-stranded break connection". Nature Reviews Genetics. 2 (3): 196–206. doi:10.1038/35056049. PMID11256071. S2CID5485937.
1 2 3 Levitt, N. C.; Hickson, I. D. (2002). "Caretaker tumour suppressor genes that defend genome integrity". Trends in Molecular Medicine. 8 (4): 179–86. doi:10.1016/s1471-4914(02)02298-0. PMID11927276.
↑ Hainut, P. 2005. ‘‘25 years of p53 research.’’ New York: Springer Publishing.
↑ Bernstein C, Bernstein H, Payne CM, Garewal H. DNA repair/pro-apoptotic dual-role proteins in five major DNA repair pathways: fail-safe protection against carcinogenesis. Mutat Res. 2002 Jun;511(2):145-78. doi: 10.1016/s1383-5742(02)00009-1. PMID: 12052432
1 2 3 4 5 6 7 Van Heemst, D; Den Reijer, P. M.; Westendorp, R. G. (2007). "Ageing or cancer: A review on the role of caretakers and gatekeepers". European Journal of Cancer. 43 (15): 2144–52. doi:10.1016/j.ejca.2007.07.011. PMID17764928.
↑ Maynard S, Fang EF, Scheibye-Knudsen M, Croteau DL, Bohr VA. DNA Damage, DNA Repair, Aging, and Neurodegeneration. Cold Spring Harb Perspect Med. 2015 Sep 18;5(10):a025130. doi: 10.1101/cshperspect.a025130. PMID: 26385091; PMCID: PMC4588127
Related Research Articles
An oncogene is a gene that has the potential to cause cancer. In tumor cells, these genes are often mutated, or expressed at high levels.
A tumor suppressor gene (TSG), or anti-oncogene, is a gene that regulates a cell during cell division and replication. If the cell grows uncontrollably, it will result in cancer. When a tumor suppressor gene is mutated, it results in a loss or reduction in its function. In combination with other genetic mutations, this could allow the cell to grow abnormally. The loss of function for these genes may be even more significant in the development of human cancers, compared to the activation of oncogenes.
DNA repair is a collection of processes by which a cell identifies and corrects damage to the DNA molecules that encode its genome. In human cells, both normal metabolic activities and environmental factors such as radiation can cause DNA damage, resulting in tens of thousands of individual molecular lesions per cell per day. Many of these lesions cause structural damage to the DNA molecule and can alter or eliminate the cell's ability to transcribe the gene that the affected DNA encodes. Other lesions induce potentially harmful mutations in the cell's genome, which affect the survival of its daughter cells after it undergoes mitosis. As a consequence, the DNA repair process is constantly active as it responds to damage in the DNA structure. When normal repair processes fail, and when cellular apoptosis does not occur, irreparable DNA damage may occur, including double-strand breaks and DNA crosslinkages. This can eventually lead to malignant tumors, or cancer as per the two hit hypothesis.
Li–Fraumeni syndrome is a rare, autosomal dominant, hereditary disorder that predisposes carriers to cancer development. It was named after two American physicians, Frederick Pei Li and Joseph F. Fraumeni, Jr., who first recognized the syndrome after reviewing the medical records and death certificates of 648 childhood rhabdomyosarcoma patients. This syndrome is also known as the sarcoma, breast, leukaemia and adrenal gland (SBLA) syndrome.
Carcinogenesis, also called oncogenesis or tumorigenesis, is the formation of a cancer, whereby normal cells are transformed into cancer cells. The process is characterized by changes at the cellular, genetic, and epigenetic levels and abnormal cell division. Cell division is a physiological process that occurs in almost all tissues and under a variety of circumstances. Normally, the balance between proliferation and programmed cell death, in the form of apoptosis, is maintained to ensure the integrity of tissues and organs. According to the prevailing accepted theory of carcinogenesis, the somatic mutation theory, mutations in DNA and epimutations that lead to cancer disrupt these orderly processes by interfering with the programming regulating the processes, upsetting the normal balance between proliferation and cell death. This results in uncontrolled cell division and the evolution of those cells by natural selection in the body. Only certain mutations lead to cancer whereas the majority of mutations do not.
Cell damage is a variety of changes of stress that a cell suffers due to external as well as internal environmental changes. Amongst other causes, this can be due to physical, chemical, infectious, biological, nutritional or immunological factors. Cell damage can be reversible or irreversible. Depending on the extent of injury, the cellular response may be adaptive and where possible, homeostasis is restored. Cell death occurs when the severity of the injury exceeds the cell's ability to repair itself. Cell death is relative to both the length of exposure to a harmful stimulus and the severity of the damage caused. Cell death may occur by necrosis or apoptosis.
Netrin receptor DCC, also known as DCC, or colorectal cancer suppressor is a protein which in humans is encoded by the DCC gene. DCC has long been implicated in colorectal cancer and its previous name was Deleted in colorectal carcinoma. Netrin receptor DCC is a single transmembrane receptor.
Inhibitor of growth protein 1 is a protein that in humans is encoded by the ING1 gene.
Deleted in Liver Cancer 1 also known as DLC1 and StAR-related lipid transfer protein 12 (STARD12) is a protein which in humans is encoded by the DLC1 gene.
Cellular senescence is a phenomenon characterized by the cessation of cell division. In their experiments during the early 1960s, Leonard Hayflick and Paul Moorhead found that normal human fetal fibroblasts in culture reach a maximum of approximately 50 cell population doublings before becoming senescent. This process is known as "replicative senescence", or the Hayflick limit. Hayflick's discovery of mortal cells paved the path for the discovery and understanding of cellular aging molecular pathways. Cellular senescence can be initiated by a wide variety of stress inducing factors. These stress factors include both environmental and internal damaging events, abnormal cellular growth, oxidative stress, autophagy factors, among many other things.
The MRN complex is a protein complex consisting of Mre11, Rad50 and Nbs1. In eukaryotes, the MRN/X complex plays an important role in the initial processing of double-strand DNA breaks prior to repair by homologous recombination or non-homologous end joining. The MRN complex binds avidly to double-strand breaks both in vitro and in vivo and may serve to tether broken ends prior to repair by non-homologous end joining or to initiate DNA end resection prior to repair by homologous recombination. The MRN complex also participates in activating the checkpoint kinase ATM in response to DNA damage. Production of short single-strand oligonucleotides by Mre11 endonuclease activity has been implicated in ATM activation by the MRN complex.
Yippee-like 3 (Drosophila) is a protein that in humans is encoded by the YPEL3 gene. YPEL3 has growth inhibitory effects in normal and tumor cell lines. One of five family members (YPEL1-5), YPEL3 was named in reference to its Drosophila melanogaster orthologue. Initially discovered in a gene expression profiling assay of p53 activated MCF7 cells, induction of YPEL3 has been shown to trigger permanent growth arrest or cellular senescence in certain human normal and tumor cell types. DNA methylation of a CpG island near the YPEL3 promoter as well as histone acetylation may represent possible epigenetic mechanisms leading to decreased gene expression in human tumors.
The hallmarks of cancer were originally six biological capabilities acquired during the multistep development of human tumors and have since been increased to eight capabilities and two enabling capabilities. The idea was coined by Douglas Hanahan and Robert Weinberg in their paper TheHallmarks of Cancer published January 2000 in Cell.
Genome instability refers to a high frequency of mutations within the genome of a cellular lineage. These mutations can include changes in nucleic acid sequences, chromosomal rearrangements or aneuploidy. Genome instability does occur in bacteria. In multicellular organisms genome instability is central to carcinogenesis, and in humans it is also a factor in some neurodegenerative diseases such as amyotrophic lateral sclerosis or the neuromuscular disease myotonic dystrophy.
Anticancer genes are genes that, when ectopically overexpressed, specifically destroy tumour cells without harming normal, untransformed cells. This cellular destruction can be due to a variety of mechanisms, such as apoptosis, mitotic catastrophe followed by apoptosis or necrosis, and autophagy. Anticancer genes emerged from studies on cancer cells in the late 1990s. Currently, there have been 291 anticancer genes discovered in the human genome. In order to be classified as an anticancer gene, the gene must have base substitutions leading to missense amino-acid changes, deletions, or insertions leading to frameshifts that alter the protein the gene codes for, increases and decreases in copy-number increases, or gene rearrangements leading to their deregulation.
Antineoplastic resistance, often used interchangeably with chemotherapy resistance, is the resistance of neoplastic (cancerous) cells, or the ability of cancer cells to survive and grow despite anti-cancer therapies. In some cases, cancers can evolve resistance to multiple drugs, called multiple drug resistance.
Chromosomal instability (CIN) is a type of genomic instability in which chromosomes are unstable, such that either whole chromosomes or parts of chromosomes are duplicated or deleted. More specifically, CIN refers to the increase in rate of addition or loss of entire chromosomes or sections of them. The unequal distribution of DNA to daughter cells upon mitosis results in a failure to maintain euploidy leading to aneuploidy. In other words, the daughter cells do not have the same number of chromosomes as the cell they originated from. Chromosomal instability is the most common form of genetic instability and cause of aneuploidy.
Judith Campisi is an American biochemist and cell biologist. She is a professor of biogerontology at the Buck Institute for Research on Aging. She is also a member of the SENS Research Foundation Advisory Board and an adviser at the Lifeboat Foundation. She is co-editor in chief of the Aging Journal, together with Mikhail Blagosklonny and David Sinclair, and founder of the pharmaceutical company Unity Biotechnology. She is listed in Who's Who in Gerontology. She is widely known for her research on how senescent cells influence aging and cancer — in particular the Senescence Associated Secretory Phenotype (SASP).
Neuroepigenetics is the study of how epigenetic changes to genes affect the nervous system. These changes may effect underlying conditions such as addiction, cognition, and neurological development.
CUX1 is an animal gene. The name stands for Cut like homeobox 1. The term "cut" derives from the "cut wing" phenotype observed in a mutant of Drosophila melanogaster. In mammals, a CCAAT-displacement activity was originally described in DNA binding assays. The human gene was identified following purification of the CCAAT-displacement protein (CDP) and has been successively been called CDP, Cut-like 1 (CUTL1), CDP/Cut and finally, CUX1.. Cut homeobox genes are present in all metazoans. In mammals, CUX1 is expressed ubiquitously in all tissues. A second gene, called CUX2, is expressed primarily in neuronal cells.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.