In some species, males will perform ritualized movements to attract females. The male six-plumed bird-of-paradise (Parotia lawesii) exemplifies male courtship display with its ritualized "ballerina dance" and unique occipital and breast feathers that serve to stimulate the female visual system.[7] In Drosophila subobscura, male courtship display is seen through the male's intricate wing scissoring patterns and rapid sidestepping.[8] These stimulations, along with many other factors, result in subsequent copulation or rejection.
In other species, males may exhibit courtship displays that serve as both visual and auditory stimulation. For example, the male Anna's hummingbird (Calypte anna) and calliope hummingbird (Stellula calliope) perform two types of courtship displays involving a combination of visual and vocal display—a stationary shuttle display[9] and dive display.[10][11] When engaging in the stationary shuttle display, the male displays a flared gorget and hovers in front of the female, moving from side to side while rotating his body and tail. The rhythmic movements of the male's wings produce a distinctive buzzing sound.[9] When conducting a dive display, the male typically ascends approximately 20–35m (66–115ft) in the air then abruptly turns and descends in a dive-like fashion. As the male flies over the female, he rotates his body and spreads his tail feathers, which flutter and collide to produce a short, buzzing sound.[9]
A male satin bowerbird makes and uses a bower to attract potential mates.
In addition, some animals attempt to attract females through the construction and decoration of unique structures. This technique can be seen in the satin bowerbird (Ptilonorhynchus violaceus) of Australia, males of which build and decorate nest-like structures called "bowers".[12] Bowers are decorated with bright and colourful objects (typically blue in colour) to attract and stimulate visiting females.[12] Typically, males who acquire the largest number of decorations tend to have greater success in mating.[13]
In some species, males initiate courtship rituals only after mounting the female. Courtship may even continue after copulation has been completed.[14] In this system, the ability of the female to choose her mate is limited. This process, known as copulatory courtship, is prevalent in many insect species.[15]
In most species, the male sex initiates courtship displays in precopulatory sexual selection. Performing a display allows the male to present his traits or abilities to a female. Mate choice, in this context, is driven by females; direct or indirect benefits to the female often determine which males reproduce and which do not.
Direct benefits may accrue to the female during male courtship displays. Females can raise their own fitness if they respond to courtship behavior that signals benefits to the female rather than the fitness of the male. For example, choosing to mate with males that produce local signals would require less energy for a female as she searches for a mate.[16] Males may compete by imposing lower mating costs on the female or even providing material or offspring contributions to the female.
Indirect benefits are benefits that may not directly affect the parents' fitness but instead increase the fitness of the offspring. Since the offspring of a female will inherit half of the genetic information from the male counterpart, those traits she saw as attractive will be passed on, producing fit offspring. In this case, males may compete during courtship by displaying desirable traits to pass on to offspring.
Female display
Female courtship display is less common in nature as a female would have to invest a lot of energy into both exaggerated traits and in their energetically expensive gametes.[17] However, situations in which males are the sexually selective sex in a species do occur in nature. Male choice in reproduction can arise if males are the sex in a species that are in short supply, for example, if there is a female bias in the operational sex ratio.[18] This could arise in mating systems where reproducing comes at an energy cost to males.[19][20] Such energy costs can include the effort associated in obtaining nuptial gifts for the female[21] or performing long courtship or copulatory behaviors.[22] An added cost from these time and energy investments may come in the form of increased male mortality rates, putting further strain on males attempting to reproduce.
In pipefish (Syngnathus typhle), females use a temporary ornament, a striped pattern, to both attract males and intimidate rival females. In this case, the female of a species developed a sexually selected signal which serves a dual function of being both attractive to mates and deterring rivals.[23]
Multi-modal signal processing
Bosc's fringe-toed lizard Acanthodactylus boskianus love bite
Many species of animals engage in some type of courtship display to attract a mate, such as dancing, the creation of sounds, and physical displays. However, many species are not limited to only one of these behaviors. The males of a species across many taxa create complex multi-component signals that have an effect on more than one sensory modality, also known as multi-modal signals.[24][25][26][27] There are two leading hypotheses about the adaptive significance of multi-modal signal processing. The multiple message hypothesis states that each signal that a male exhibits will contribute to a possible mate's perception of the male. The redundant signal hypothesis states that the male exhibits multiple signals that portray the same "message" to the female, with each extra signal acting as a fall-back plan for the male should there be a signaling error.[28] The choosy sex may only evaluate one, or a couple, of traits at a given time when interpreting complex signals from the opposite sex. Alternatively, the choosy sex may attempt to process all of the signals at once to facilitate evaluation of the opposite sex.[29][30]
The process of multi-modal signaling is believed to help facilitate the courtship process in many species. One such species in which multi-modal signaling is seen to improve mating success is the green tree frog (Hyla cinerea). Many anuran amphibians, such as the green tree frog, may use visual cues as well as auditory signals to increase their chances of impressing a mate.[25] When the calls of the tree frogs were held equal, it was determined that females tended to overlook an auditory-only stimulus in favor of males who combined auditory/visual multi-modal signals. It was seen that female green tree frogs preferred when males coupled the visual display with the auditory communication, concluding that male green tree frogs that are visually accessible can increase their probability of mating success.[25]
Peacock spiders (Maratus volans) are exceptionally sexually dimorphic in appearance and signaling behavior. During courtship, male peacock spiders compete using both visual displays and vibratory signals for intersexual communication.[26] Because of the intense sexual selection on male peacock spiders, the reproductive success of an individual relies heavily on a male spider's ability to combine visual and vibratory displays during courtship. The combination of these displays in courtship offers support both to the redundant signal and multiple messages hypotheses for the evolution of multi-modal signaling in species.[31]
Multi-modal signaling is not limited to males. Females in certain species have more than one trait or characteristic that they use in a courtship display to attract mates. In dance flies (Rhamphomyia longicauda), females have two ornaments — inflatable abdominal sacs and pinnate tibial scales — that they use as courtship displays in mating swarms. Intermediate variations of such female-specific ornaments are sexually selected for by male dance flies in wild populations. These ornaments may also be a signal of high fecundity in females.[32]
Mutual display
Male and female long-billed curlew, Numenius americanus, mutual courtship display
Often, males and females will perform synchronized or responsive courtship displays in a mutual fashion. With many socially monogamous species such as birds, their duet facilitates pre-copulatory reassurance of pair bonding and strengthens post-copulatory dedication to the development of offspring (e.g., great crested grebe, Podiceps cristatus).[33] For example, male and female crested auklets, Aethia cristatella, will cackle at one another as a vocal form of mutual display that serves to strengthen a bond between the two.[34] In some cases, males may pair up to perform mutual, cooperative displays in order to increase courtship success and attract females. This phenomenon can be seen with long-tailed manakins, Chiroxiphia linearis.[35]
Wild turkeys (Meleagris gallopavo) also engage in co-operative displays in which small groups of males (typically brothers) work together to attract females and deter other competitive males.[36] In many cases, only one male within the group will mate, typically the dominant male.[37] To explain this behaviour, Hamilton's theory of kin selection suggests that subordinate males receive indirect benefits by helping related males copulate successfully.[38]
Sexual ornaments
Sexual ornaments can serve to increase attractiveness[39] and indicate good genes and higher levels of fitness.[40] When exposed to exaggerated male traits, some females may respond by increasing maternal investments. For example, female canaries have been shown to produce larger and denser eggs in response to male supranormal song production.[41]
Sexual conflict
Sexual conflict is the phenomenon in which the interests of males and females in reproduction are not the same: they are often quite different:[42]
Males: their interest is to mate with a large number of completely faithful females, thus spreading their genes widely throughout a population.
Females: their interest is to mate with a large number of fit males, thus producing a large quantity of fit and varied offspring.
This has many consequences. Courtship displays allow the mate performing the selection to have a means on which to base the copulatory decision. If a female chooses more than one male, then sperm competition comes into play. This is competition between sperm to fertilize an egg, which is very competitive as only a single sperm will achieve union.[43] In some insects, the male injects a cocktail of chemicals in seminal fluid together with sperm. The chemicals kill off older sperm from any previous mates, up-regulates the female's egg-laying rate, and reduces her desire to re-mate with another male. The cocktail also shortens the female's lifespan, also reducing her likelihood of mating with other males.[44] Also, some females can get rid of the previous male's sperm.[45]
After mating has taken place, males perform various actions to prevent females from mating again. What action is performed depends on the animal. In some species, the male produces a mating plug after insemination. In some hymenoptera, the male provides a huge quantity of sperm, enough to last the female's entire life. In some birds and mammals, the male may participate in agonistic behaviors with other candidate males.[42][46][47]
Agonistic behavior and courtship
Although rare, agonistic behavior between males and females during courtship displays is seen in nature. Intraspecific agonistic behavior that results in the death of a combatant is rare because of the associated risk of death or injury. However, agonistic behavior that turns dangerous does occur.
In some species, physical traits that are sexually selected for in male courtship displays may also be used in agonistic behavior between two males for a mate. In fiddler crabs (genus Uca), males have been sexually selected to have one enlarged claw, which can take up anywhere from a third to a half of their total body mass, and one regular claw. Although the enlarged claw is believed to have developed for use in combat for territorial defense, it is not uncommon for males to employ this claw in battle for a mate.[48] Even though this claw developed as a weapon, it is also closely linked with the crabs' courtship display: it is waved in a certain pattern to attract females for mating.[49]
Agonistic behavior in courtship displays is not limited to male-male interactions. In many primate species, males direct agonistic behavior toward females prior to courtship behaviors. Such behavior can include aggressive vocalizations, displays, and physical aggression.[50] In the western gorilla (Gorilla gorilla), dominant males exhibit agonistic behavior toward female gorillas at very high rates, with the majority of those interactions being courtship-related. Most documented cases of male gorilla aggression toward females is courtship related and is used primarily as a strategy to prevent females from migrating to another male.[51]
In many cases, male courtship displays will cause forms of contest competition to develop. This is often seen within lek mating systems. For example, males will seek to obtain a certain spot or position to perform their courtship display. The best spots are regions of high contention as many males want them for themselves. Because of this direct conflict, agonistic[clarification needed] encounters between males are fairly common.[citation needed]
Extended courtship period
Mating is preceded by a courtship/pairing period in many animal mating systems. It is during this period that sexually mature animals select their partners for reproduction.[52] This courtship period, which involves displays to attract a mate by a member of a species, is usually short, lasting anywhere from 15 minutes to a few days. However, certain animals may undergo an extended courtship period, lasting as long as two months.[53]
One such exception is the emperor penguin (Aptenodytes forsteri). Emperor penguins engage in an extended courtship period that can last up to two months, the longest of any Arctic seabird. Their courtship period accounts for 16% of the total time they spend breeding, whereas in their closest relatives, the king penguin (Aptenodytes patagonicus), the courtship period takes up just three per cent of their breeding cycle.[53][54]
Energetic costs
Greater sage-grouse lek males perform a courtship "strutting" display for visiting females.
Courtship displays typically involve some sort of metabolic cost to the animal performing it.[11] The energy expended to perform courtship behaviour can vary among species. Some animals engage in displays that expend little energy, as seen in the salamander (Desmognathus ochrophaeus).[55] Under laboratory settings, courtship behaviours in this species, although complex and involving the release of pheromones,[56] represent as little as approximately one per cent of its daily calorie intake.[55]
In contrast, species that engage in prolonged or elaborate displays expend considerable amounts of energy and run the risk of developing fatigue. To prepare and prevent such a risk, some animals may gain weight before a courtship period, only to lose the weight afterward. An example of this can be seen in the greater sage-grouse (Centrocercus urophasianus). During the peak of their breeding season, which lasts up to three months during spring,[57] leks are frequently visited by groups of up to seventy females.[58] In response to such a large presence of females, males engage in a strutting display up to six to ten times per minute[59][60] for approximately three to four hours per day.[58] This frequent and repetitive behaviour can result in energy expenditures of up to 2524 kJ/day compared to the inactive males that typically expend 1218 kJ/day.[11]
Environmental factors
Various environmental factors, such as temperature, photoperiod, resource and light availability, have an effect on the timing and effectiveness of courtship displays in certain species of animals.[53]
In guppies (Poecilia reticulata), variation in the light environment plays a huge role in their ability to attract mates.[61] Guppy males alter both their 'courtship mode', whether they perform a full courtship display or try to 'engage' in sneak copulations, and distance from females as light intensity changes.[62] Courtship mode also varies with light spectrum and relates to predation risk.[63] On average, male guppies seek out and spend more time in the environment in which their color pattern is the most visible. Males, in the light environment that made them most visible, copulated with the most females.[61]
In emperor penguins (Aptenodytes forsteri), resource availability determines when male emperor penguins will be able to return to their breeding grounds to initiate their courtship rituals.[53] The greater the concentration of resources in their feeding ground, the quicker they will be able to restore their body reserves for winter, and the sooner they will be able to return to their breeding grounds. An early return to their breeding grounds comes with an increased likelihood of finding a mate.[64]
The effectiveness of Hirtodrosophila mycetophaga mating displays is influenced by the color of the bracket fungus that it mates and courts upon;[65] these flies choose brackets that are lighter, making their displays more visible to the opposite sex.[65]
Evolutionary significance
Male and female Pigeon Guillemot, Cepphus columba, mutual courtship display
There are multiple hypotheses about how courtship displays may have evolved in animals, including the Fisherian runaway model and the good genes hypothesis.
As explained by the Fisherian runaway model, sexually dimorphic males with exaggerated ornamentation may have been sexually selected for in species with female choice. Fitness of these males would increase, resulting in the proliferation of males with such ornamentation over time.[66] This means that a gene or set of genes will be favoured by female choice over time. This would explain why and how such elaborate traits develop within certain species. However, as time goes on and generations pass, the survival advantage associated with one trait may dissipate due to extreme exaggeration to the point that it decreases fitness.
The "good genes" hypothesis proposes that female selection of a mate is dependent on whether or not the male has genes that would increase the quality of the offspring of the female. In some cases, exaggerated male ornamentation may be indicative to a choosing female that a male who is able to place such a large investment in a trait somewhat counterintuitive to survival would carry good genes.[67] For example, the costs associated with bright and complex plumage can be high. Only males with good genes are able to support a large investment into the development of such traits, which, in turn displays their high fitness.
An alternative is the sensory exploitation hypothesis, which supposes that sexual preferences are the result of preexisting sensory biases, such as that for supernormal stimuli. These could drive the evolution of courtship displays.[68]
Sexual selection is a mode of natural selection in which members of one biological sex choose mates of the other sex to mate with, and compete with members of the same sex for access to members of the opposite sex. These two forms of selection mean that some individuals have greater reproductive success than others within a population, for example because they are more attractive or prefer more attractive partners to produce offspring. Successful males benefit from frequent mating and monopolizing access to one or more fertile females. Females can maximise the return on the energy they invest in reproduction by selecting and mating with the best males.
A lek is an aggregation of male animals gathered to engage in competitive displays and courtship rituals, known as lekking, to entice visiting females which are surveying prospective partners with which to mate. A lek can also indicate an available plot of space able to be utilized by displaying males to defend their own share of territory for the breeding season. A lekking species is characterised by male displays, strong female mate choice, and the conferring of indirect benefits to males and reduced costs to females. Although most prevalent among birds such as black grouse, lekking is also found in a wide range of vertebrates including some bony fish, amphibians, reptiles, and mammals, and arthropods including crustaceans and insects.
Behavioral ecology, also spelled behavioural ecology, is the study of the evolutionary basis for animal behavior due to ecological pressures. Behavioral ecology emerged from ethology after Niko Tinbergen outlined four questions to address when studying animal behaviors: What are the proximate causes, ontogeny, survival value, and phylogeny of a behavior?
Sperm competition is the competitive process between spermatozoa of two or more different males to fertilize the same egg during sexual reproduction. Competition can occur when females have multiple potential mating partners. Greater choice and variety of mates increases a female's chance to produce more viable offspring. However, multiple mates for a female means each individual male has decreased chances of producing offspring. Sperm competition is an evolutionary pressure on males, and has led to the development of adaptations to increase male's chance of reproductive success. Sperm competition results in a sexual conflict between males and females. Males have evolved several defensive tactics including: mate-guarding, mating plugs, and releasing toxic seminal substances to reduce female re-mating tendencies to cope with sperm competition. Offensive tactics of sperm competition involve direct interference by one male on the reproductive success of another male, for instance by physically removing another male's sperm prior to mating with a female. For an example, see Gryllus bimaculatus.
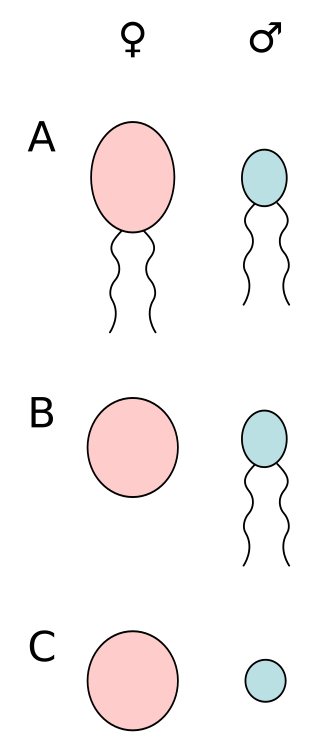
Anisogamy is a form of sexual reproduction that involves the union or fusion of two gametes that differ in size and/or form. The smaller gamete is male, a sperm cell, whereas the larger gamete is female, typically an egg cell. Anisogamy is predominant among multicellular organisms. In both plants and animals gamete size difference is the fundamental difference between females and males.
Monogamous pairing in animals refers to the natural history of mating systems in which species pair bond to raise offspring. This is associated, usually implicitly, with sexual monogamy.
Display behaviour is a set of ritualized behaviours that enable an animal to communicate to other animals about specific stimuli. Such ritualized behaviours can be visual, but many animals depend on a mixture of visual, audio, tactical and chemical signals. Evolution has tailored these stereotyped behaviours to allow animals to communicate both conspecifically and interspecifically which allows for a broader connection in different niches in an ecosystem. It is connected to sexual selection and survival of the species in various ways. Typically, display behaviour is used for courtship between two animals and to signal to the female that a viable male is ready to mate. In other instances, species may make territorial displays, in order to preserve a foraging or hunting territory for its family or group. A third form is exhibited by tournament species in which males will fight in order to gain the 'right' to breed. Animals from a broad range of evolutionary hierarchies avail of display behaviours - from invertebrates such as the simple jumping spider to the more complex vertebrates like the harbour seal.
Sexual conflict or sexual antagonism occurs when the two sexes have conflicting optimal fitness strategies concerning reproduction, particularly over the mode and frequency of mating, potentially leading to an evolutionary arms race between males and females. In one example, males may benefit from multiple matings, while multiple matings may harm or endanger females, due to the anatomical differences of that species. Sexual conflict underlies the evolutionary distinction between male and female.
Mate choice is one of the primary mechanisms under which evolution can occur. It is characterized by a "selective response by animals to particular stimuli" which can be observed as behavior. In other words, before an animal engages with a potential mate, they first evaluate various aspects of that mate which are indicative of quality—such as the resources or phenotypes they have—and evaluate whether or not those particular trait(s) are somehow beneficial to them. The evaluation will then incur a response of some sort.
Sexual cannibalism is when an animal, usually the female, cannibalizes its mate prior to, during, or after copulation. It is a trait observed in many arachnid orders and several insect and crustacean clades. Several hypotheses to explain this seemingly paradoxical behavior have been proposed. The adaptive foraging hypothesis, aggressive spillover hypothesis and mistaken identity hypothesis are among the proposed hypotheses to explain how sexual cannibalism evolved. This behavior is believed to have evolved as a manifestation of sexual conflict, occurring when the reproductive interests of males and females differ. In many species that exhibit sexual cannibalism, the female consumes the male upon detection. Females of cannibalistic species are generally hostile and unwilling to mate; thus many males of these species have developed adaptive behaviors to counteract female aggression.
Many species have multiple sexual ornaments, whereby females select mating partners using several cues instead of only one cue. Whereas this phenomenon is self-evident and hence long recognized, adaptive explanations of why females use several instead of only one signal have been formulated relatively recently. Several hypotheses exist, but mutually exclusive tests are still lacking.
A biological ornament is a characteristic of an animal that appears to serve a decorative function rather than a utilitarian function. Many are secondary sexual characteristics, and others appear on young birds during the period when they are dependent on being fed by their parents. Ornaments are used in displays to attract mates, which may lead to the evolutionary process known as sexual selection. An animal may shake, lengthen, or spread out its ornament in order to get the attention of the opposite sex, which will in turn choose the most attractive one with which to mate. Ornaments are most often observed in males, and choosing an extravagantly ornamented male benefits females as the genes that produce the ornament will be passed on to her offspring, increasing their own reproductive fitness. As Ronald Fisher noted, the male offspring will inherit the ornament while the female offspring will inherit the preference for said ornament, which can lead to a positive feedback loop known as a Fisherian runaway. These structures serve as cues to animal sexual behaviour, that is, they are sensory signals that affect mating responses. Therefore, ornamental traits are often selected by mate choice.
Interlocus sexual conflict is a type of sexual conflict that occurs through the interaction of a set of antagonistic alleles at two or more different loci, or the location of a gene on a chromosome, in males and females, resulting in the deviation of either or both sexes from the fitness optima for the traits. A co-evolutionary arms race is established between the sexes in which either sex evolves a set of antagonistic adaptations that is detrimental to the fitness of the other sex. The potential for reproductive success in one organism is strengthened while the fitness of the opposite sex is weakened. Interlocus sexual conflict can arise due to aspects of male–female interactions such as mating frequency, fertilization, relative parental effort, female remating behavior, and female reproductive rate.
Female copulatory vocalizations, also called female copulation calls or coital vocalizations, are produced by female primates, including human females, and female non-primates. Copulatory vocalizations usually occur during copulation and are hence related to sexual activity. Vocalizations that occur before intercourse, for the purpose of attracting mates, are known as mating calls.
Sexual selection in birds concerns how birds have evolved a variety of mating behaviors, with the peacock tail being perhaps the most famous example of sexual selection and the Fisherian runaway. Commonly occurring sexual dimorphisms such as size and color differences are energetically costly attributes that signal competitive breeding situations. Many types of avian sexual selection have been identified; intersexual selection, also known as female choice; and intrasexual competition, where individuals of the more abundant sex compete with each other for the privilege to mate. Sexually selected traits often evolve to become more pronounced in competitive breeding situations until the trait begins to limit the individual's fitness. Conflicts between an individual fitness and signaling adaptations ensure that sexually selected ornaments such as plumage coloration and courtship behavior are "honest" traits. Signals must be costly to ensure that only good-quality individuals can present these exaggerated sexual ornaments and behaviors.
Sexual selection in mammals is a process the study of which started with Charles Darwin's observations concerning sexual selection, including sexual selection in humans, and in other mammals, consisting of male–male competition and mate choice that mold the development of future phenotypes in a population for a given species.
Sexual selection in scaled reptiles studies how sexual selection manifests in snakes and lizards, which constitute the order Squamata of reptiles. Each of the over three thousand snakes use different tactics in acquiring mates. Ritual combat between males for the females they want to mate with includes topping, a behavior exhibited by most viperids in which one male will twist around the vertically elevated fore body of its opponent and forcing it downward. It is common for neck biting to occur while the snakes are entwined.
Sexual selection in amphibians involves sexual selection processes in amphibians, including frogs, salamanders and newts. Prolonged breeders, the majority of frog species, have breeding seasons at regular intervals where male-male competition occurs with males arriving at the waters edge first in large number and producing a wide range of vocalizations, with variations in depth of calls the speed of calls and other complex behaviours to attract mates. The fittest males will have the deepest croaks and the best territories, with females making their mate choices at least partly based on the males depth of croaking. This has led to sexual dimorphism, with females being larger than males in 90% of species, males in 10% and males fighting for groups of females.
Sexual selection in spiders shows how sexual selection explains the evolution of phenotypic traits in spiders. Male spiders have many complex courtship rituals and have to avoid being eaten by the females, with the males of most species survive a few matings, and having short life spans.
The sensory trap hypothesis describes an evolutionary idea that revolves around mating behavior and female mate choice. It is a model of female preference and male sexual trait evolution through what is known as sensory exploitation. Sensory exploitation, or a sensory trap is an event that occurs in nature where male members of a species perform behaviors or display visual traits that resemble a non-sexual stimulus which females are responsive to. This tricks females into engaging with the males, thus creating more mating opportunities for males. What makes it a sensory trap is that these female responses evolved in a non-sexual context, and the male produced stimulus exploits the female response which would not otherwise occur without the mimicked stimulus.
↑ Riede, Tobias; Forstmeier, Wolfgang; Kempenaers, Bart; Goller, Franz (2015). "The functional morphology of male courtship displays in the Pectoral Sandpiper (Calidris melanotos)". The Auk. 132: 65–77. doi:10.1642/AUK-14-25.1. S2CID86155377.
1 2 3 Clark, Christopher J. (2012). "The role of power versus energy in courtship: what is the 'energetic cost' of a courtship display?". Animal Behaviour. 84 (1): 269–277. doi:10.1016/j.anbehav.2012.04.012. S2CID45711419.
1 2 Borgia, Gerald (1985). "Bower quality, number of decorations and mating success of male satin bowerbirds (Ptilonorhynchus violaceus): an experimental analysis". Animal Behaviour. 33 (1): 266–271. doi:10.1016/s0003-3472(85)80140-8. S2CID53192968.
↑ Madden, Joah R. (2003). "Male spotted bowerbirds preferentially choose, arrange and proffer objects that are good predictors of mating success". Behavioral Ecology and Sociobiology. 53 (5): 263–268. doi:10.1007/s00265-003-0582-7. JSTOR4602214. S2CID44048073.
↑ Simmons, L.W. (July 1990). "Nuptial feeding in tettigoniids male costs and the rates of fecundity increase". Behavioral Ecology and Sociobiology. 27 (1): 43–47. doi:10.1007/BF00183312. JSTOR4600442. S2CID24088701.
↑ Saeki, Yoriko; Kruse, Kipp C.; Switzer, Paul V. (September 2005). "Physiological costs of mate guarding in the Japanese beetle (Popillia japonica Newman)". Ethology. 111 (9): 863–877. Bibcode:2005Ethol.111..863S. doi:10.1111/j.1439-0310.2005.01106.x.
1 2 3 Laird, Krispen L.; Clements, Paul; Hunter, Kimberly L.; Taylor, Ryan C. (2016). "Multimodal signaling improves mating success in the green tree frog (Hyla cinerea), but may not help small males". Behavioral Ecology and Sociobiology. 70 (9): 1517–1525. doi:10.1007/s00265-016-2160-9. S2CID15612885.
↑ Zubakin, V. A.; Volodin, I. A.; Klenova, A. V.; Zubakina, E. V.; Volodina, E. V.; Lapshina, E. N. (2010). "Behavior of crested auklets (Aethia cristatella, Charadriiformes, Alcidae) in the breeding season: Visual and acoustic displays". Biology Bulletin. 37 (8): 823–835. Bibcode:2010BioBu..37..823Z. doi:10.1134/S1062359010080066. S2CID9709011.
↑ Lukianchuk, K. C.; Doucet, S. M. (2014). "Cooperative courtship display in Long-tailed Manakins Chiroxiphia linearis: predictors of courtship success revealed through full characterization of display". Journal of Ornithology. 155 (3): 729–743. doi:10.1007/s10336-014-1059-3. S2CID15592813.
↑ Tibbetts, Elizabeth A.; Forrest, Taylor; Vernier, Cassondra; Jinn, Judy; Madagame, Andrew (2015-11-01). "Socially selected ornaments and fitness: Signals of fighting ability in paper wasps are positively associated with survival, reproductive success, and rank". Evolution. 69 (11): 2917–2926. doi:10.1111/evo.12793. hdl:2027.42/134447. ISSN1558-5646. PMID26456701. S2CID10079068.
↑ Garcia-Fernandez, V.; Draganoiu, T.I.; Ung, D.; Lacroix, A.; Malacarne, G.; Leboucher, G. (2013). "Female canaries invest more in response to an exaggerated male trait". Animal Behaviour. 85 (3): 679–684. doi:10.1016/j.anbehav.2013.01.007. S2CID53154422.
1 2 Arnqvist, G. & Rowe, L. 2005. Sexual conflict. Princeton, New Jersey. Princeton University Press.
↑ Kahn, A. T. (2014). "Female preferences for timing in a fiddler crab with synchronous courtship waving displays". Animal Behaviour. 98: 35–39. doi:10.1016/j.anbehav.2014.09.028. S2CID44139315.
↑ Muller, M. N. (2009). Sexual Coercion in Primates and Humans. Harvard University Press. pp.3–22. ISBN9780674033245.
↑ Isenmann, P. (1971). "Contribution a' l'e´thologie et a' l'e´cologie du manchot empereur (Aptenodytes forsteri Gray) a' la colonie de Pointe Ge´ologie (Terre Ade´lie)". L'Oiseau et la RFO. 40: 136–159.
↑ Houck, Lynne D.; Arnold, Stevan J.; Thisted, Ronald A. (1985). "A Statistical Study of Mate Choice: Sexual Selection in a Plethodontid Salamander (Desmognathus ochrophaeus)". Evolution. 39 (2): 370–386. doi:10.2307/2408370. JSTOR2408370. PMID28564228.
↑ Vehrencamp, Sandra L.; Bradbury, Jack W.; Gibson, Robert M. (1989). "The energetic cost of display in male sage grouse". Animal Behaviour. 38 (5): 885–896. doi:10.1016/s0003-3472(89)80120-4. S2CID53195569.
↑ Endler, J. A. (2016). "Predation, light intensity and courtship behaviour in Poecilia reticulata (Pisces: Poeciliidae)". Animal Behaviour. 35 (5): 1376–1385. doi:10.1016/S0003-3472(87)80010-6. S2CID53169408.
↑ Gamble, S. (2003). "Environmental variation and the maintenance of polymorphism: the effect of ambient light spectrum on mating behaviour and sexual selection in guppies". Ecology Letters. 6 (5): 463–472. Bibcode:2003EcolL...6..463G. doi:10.1046/j.1461-0248.2003.00449.x.
↑ Yasui, Y. (1997). "A "Good-Sperm" model can explain the evolution of costly multiple mating by females". The American Naturalist. 149 (3): 573. doi:10.1086/286006. S2CID84991216.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.