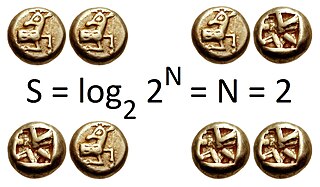A diversity index is a quantitative measure that reflects how many different types (e.g. species) there are in a dataset (e.g. a community). More sophisticated indices accounting for the phylogenetic relatedness among the types.[1] Diversity indices are statistical representations of different aspects of biodiversity (e.g. richness, evenness, and dominance), that are useful simplifications to compare different communities or sites.
When diversity indices are used in ecology, the types of interest are usually species, but they can also be other categories, such as genera, families, functional types, or haplotypes. The entities of interest are usually individual organisms (e.g. plants or animals), and the measure of abundance can be, for example, number of individuals, biomass or coverage. In demography, the entities of interest can be people, and the types of interest various demographic groups. In information science, the entities can be characters and the types of the different letters of the alphabet. The most commonly used diversity indices are simple transformations of the effective number of types (also known as 'true diversity'), but each diversity index can also be interpreted in its own right as a measure corresponding to some real phenomenon (but a different one for each diversity index).[2][3][4][5]
Many indices only account for categorical diversity between subjects or entities. Such indices, however do not account for the total variation (diversity) that can be held between subjects or entities which occurs only when both categorical and qualitative diversity are calculated.
True diversity, or the effective number of types, refers to the number of equally abundant types needed for the average proportional abundance of the types to equal that observed in the dataset of interest (where all types may not be equally abundant). The true diversity in a dataset is calculated by first taking the weighted generalized meanMq−1 of the proportional abundances of the types in the dataset, and then taking the reciprocal of this. The equation is:[4][5]
The denominatorMq−1 equals the average proportional abundance of the types in the dataset as calculated with the weighted generalized mean with exponent q−1. In the equation, R is richness (the total number of types in the dataset), and the proportional abundance of the ith type is pi. The proportional abundances themselves are used as the nominal weights. The numbers are called Hill numbers of orderq or effective number of species.[6]
When q = 1, the above equation is undefined. However, the mathematical limit as q approaches 1 is well defined and the corresponding diversity is calculated with the following equation:
which is the exponential of the Shannon entropy calculated with natural logarithms (see above). In other domains, this statistic is also known as the perplexity.
The general equation of diversity is often written in the form[2][3]
and the term inside the parentheses is called the basic sum. Some popular diversity indices correspond to the basic sum as calculated with different values of q.[3]
Sensitivity of the diversity value to rare vs. abundant species
The value of q is often referred to as the order of the diversity. It defines the sensitivity of the true diversity to rare vs. abundant species by modifying how the weighted mean of the species' proportional abundances is calculated. With some values of the parameter q, the value of the generalized mean Mq−1 assumes familiar kinds of weighted means as special cases. In particular,
As q approaches infinity, the weighted generalized mean with exponent q−1 approaches the maximum pi value, which is the proportional abundance of the most abundant species in the dataset.
Generally, increasing the value of q increases the effective weight given to the most abundant species. This leads to obtaining a larger Mq−1 value and a smaller true diversity (qD) value with increasing q.
When q = 1, the weighted geometric mean of the pi values is used, and each species is exactly weighted by its proportional abundance (in the weighted geometric mean, the weights are the exponents). When q > 1, the weight given to abundant species is exaggerated, and when q < 1, the weight given to rare species is. At q = 0, the species weights exactly cancel out the species proportional abundances, such that the weighted mean of the pi values equals 1 / R even when all species are not equally abundant. At q = 0, the effective number of species, 0D, hence equals the actual number of species R. In the context of diversity, q is generally limited to non-negative values. This is because negative values of q would give rare species so much more weight than abundant ones that qD would exceed R.[4][5]
Richness R simply quantifies how many different types the dataset of interest contains. For example, species richness (usually noted S) is simply the number of species, e.g. at a particular site. Richness is a simple measure, so it has been a popular diversity index in ecology, where abundance data are often not available.[7] If true diversity is calculated with q = 0, the effective number of types (0D) equals the actual number of types, which is identical to Richness (R).[3][5]
Shannon index
The Shannon index has been a popular diversity index in the ecological literature, where it is also known as Shannon's diversity index, Shannon–Wiener index, and (erroneously) Shannon–Weaver index.[8] The measure was originally proposed by Claude Shannon in 1948 to quantify the entropy (hence Shannon entropy, related to Shannon information content) in strings of text.[9] The idea is that the more letters there are, and the closer their proportional abundances in the string of interest, the more difficult it is to correctly predict which letter will be the next one in the string. The Shannon entropy quantifies the uncertainty (entropy or degree of surprise) associated with this prediction. It is most often calculated as follows:
where pi is the proportion of characters belonging to the ith type of letter in the string of interest. In ecology, pi is often the proportion of individuals belonging to the ith species in the dataset of interest. Then the Shannon entropy quantifies the uncertainty in predicting the species identity of an individual that is taken at random from the dataset.
Although the equation is here written with natural logarithms, the base of the logarithm used when calculating the Shannon entropy can be chosen freely. Shannon himself discussed logarithm bases 2, 10 and e, and these have since become the most popular bases in applications that use the Shannon entropy. Each log base corresponds to a different measurement unit, which has been called binary digits (bits), decimal digits (decits), and natural digits (nats) for the bases 2, 10 and e, respectively. Comparing Shannon entropy values that were originally calculated with different log bases requires converting them to the same log base: change from the base a to base b is obtained with multiplication by logba.[9]
The Shannon index (H') is related to the weighted geometric mean of the proportional abundances of the types. Specifically, it equals the logarithm of true diversity as calculated with q = 1:[4]
This can also be written
which equals
Since the sum of the pi values equals 1 by definition, the denominator equals the weighted geometric mean of the pi values, with the pi values themselves being used as the weights (exponents in the equation). The term within the parentheses hence equals true diversity 1D, and H' equals ln(1D).[2][4][5]
When all types in the dataset of interest are equally common, all pi values equal 1 / R, and the Shannon index hence takes the value ln(R). The more unequal the abundances of the types, the larger the weighted geometric mean of the pi values, and the smaller the corresponding Shannon entropy. If practically all abundance is concentrated to one type, and the other types are very rare (even if there are many of them), Shannon entropy approaches zero. When there is only one type in the dataset, Shannon entropy exactly equals zero (there is no uncertainty in predicting the type of the next randomly chosen entity).
In machine learning the Shannon index is also called as Information gain.
Rényi entropy
The Rényi entropy is a generalization of the Shannon entropy to other values of q than 1. It can be expressed:
which equals
This means that taking the logarithm of true diversity based on any value of q gives the Rényi entropy corresponding to the same value of q.
Simpson index
The Simpson index was introduced in 1949 by Edward H. Simpson to measure the degree of concentration when individuals are classified into types.[10] The same index was rediscovered by Orris C. Herfindahl in 1950.[11] The square root of the index had already been introduced in 1945 by the economist Albert O. Hirschman.[12] As a result, the same measure is usually known as the Simpson index in ecology, and as the Herfindahl index or the Herfindahl–Hirschman index (HHI) in economics.
The measure equals the probability that two entities taken at random from the dataset of interest represent the same type.[10] It equals:
where R is richness (the total number of types in the dataset). This equation is also equal to the weighted arithmetic mean of the proportional abundances pi of the types of interest, with the proportional abundances themselves being used as the weights.[2] Proportional abundances are by definition constrained to values between zero and one, but it is a weighted arithmetic mean, hence λ ≥ 1/R, which is reached when all types are equally abundant.
By comparing the equation used to calculate λ with the equations used to calculate true diversity, it can be seen that 1/λ equals 2D, i.e., true diversity as calculated with q = 2. The original Simpson's index hence equals the corresponding basic sum.[3]
The interpretation of λ as the probability that two entities taken at random from the dataset of interest represent the same type assumes that the first entity is replaced to the dataset before taking the second entity. If the dataset is very large, sampling without replacement gives approximately the same result, but in small datasets, the difference can be substantial. If the dataset is small, and sampling without replacement is assumed, the probability of obtaining the same type with both random draws is:
where ni is the number of entities belonging to the ith type and N is the total number of entities in the dataset.[10] This form of the Simpson index is also known as the Hunter–Gaston index in microbiology.[13]
Since the mean proportional abundance of the types increases with decreasing number of types and increasing abundance of the most abundant type, λ obtains small values in datasets of high diversity and large values in datasets of low diversity. This is counterintuitive behavior for a diversity index, so often, such transformations of λ that increase with increasing diversity have been used instead. The most popular of such indices have been the inverse Simpson index (1/λ) and the Gini–Simpson index (1−λ).[2][3] Both of these have also been called the Simpson index in the ecological literature, so care is needed to avoid accidentally comparing the different indices as if they were the same.
Inverse Simpson index
The inverse Simpson index equals:
This simply equals true diversity of order 2, i.e. the effective number of types that is obtained when the weighted arithmetic mean is used to quantify average proportional abundance of types in the dataset of interest.
The Gini-Simpson Index is also called Gini impurity, or Gini's diversity index[14] in the field of Machine Learning. The original Simpson index λ equals the probability that two entities taken at random from the dataset of interest (with replacement) represent the same type. Its transformation 1−λ, therefore, equals the probability that the two entities represent different types. This measure is also known in ecology as the probability of interspecific encounter (PIE)[15] and the Gini–Simpson index.[3] It can be expressed as a transformation of the true diversity of order 2:
The Gibbs–Martin index of sociology, psychology, and management studies,[16] which is also known as the Blau index, is the same measure as the Gini–Simpson index.
The Berger–Parker[17] index equals the maximum pi value in the dataset, i.e., the proportional abundance of the most abundant type. This corresponds to the weighted generalized mean of the pi values when q approaches infinity, and hence equals the inverse of the true diversity of order infinity (1/∞D).
In mathematics, the geometric mean is a mean or average which indicates a central tendency of a finite set of real numbers by using the product of their values. The geometric mean is defined as the nth root of the product of n numbers, i.e., for a set of numbers a1, a2, ..., an, the geometric mean is defined as
In information theory, the entropy of a random variable is the average level of "information", "surprise", or "uncertainty" inherent to the variable's possible outcomes. Given a discrete random variable , which takes values in the alphabet and is distributed according to , the entropy is
In probability theory and statistics, the Weibull distribution is a continuous probability distribution. It models a broad range of random variables, largely in the nature of a time to failure or time between events. Examples are maximum one-day rainfalls and the time a user spends on a web page.
In physics, a partition function describes the statistical properties of a system in thermodynamic equilibrium. Partition functions are functions of the thermodynamic state variables, such as the temperature and volume. Most of the aggregate thermodynamic variables of the system, such as the total energy, free energy, entropy, and pressure, can be expressed in terms of the partition function or its derivatives. The partition function is dimensionless.
Species diversity is the number of different species that are represented in a given community. The effective number of species refers to the number of equally abundant species needed to obtain the same mean proportional species abundance as that observed in the dataset of interest. Meanings of species diversity may include species richness, taxonomic or phylogenetic diversity, and/or species evenness. Species richness is a simple count of species. Taxonomic or phylogenetic diversity is the genetic relationship between different groups of species. Species evenness quantifies how equal the abundances of the species are.
In mathematical statistics, the Kullback–Leibler (KL) divergence, denoted , is a type of statistical distance: a measure of how one probability distribution P is different from a second, reference probability distribution Q. A simple interpretation of the KL divergence of P from Q is the expected excess surprise from using Q as a model when the actual distribution is P. While it is a measure of how different two distributions are, and in some sense is thus a "distance", it is not actually a metric, which is the most familiar and formal type of distance. In particular, it is not symmetric in the two distributions, and does not satisfy the triangle inequality. Instead, in terms of information geometry, it is a type of divergence, a generalization of squared distance, and for certain classes of distributions, it satisfies a generalized Pythagorean theorem.
Variational Bayesian methods are a family of techniques for approximating intractable integrals arising in Bayesian inference and machine learning. They are typically used in complex statistical models consisting of observed variables as well as unknown parameters and latent variables, with various sorts of relationships among the three types of random variables, as might be described by a graphical model. As typical in Bayesian inference, the parameters and latent variables are grouped together as "unobserved variables". Variational Bayesian methods are primarily used for two purposes:
To provide an analytical approximation to the posterior probability of the unobserved variables, in order to do statistical inference over these variables.
To derive a lower bound for the marginal likelihood of the observed data. This is typically used for performing model selection, the general idea being that a higher marginal likelihood for a given model indicates a better fit of the data by that model and hence a greater probability that the model in question was the one that generated the data.
In information theory, the Rényi entropy is a quantity that generalizes various notions of entropy, including Hartley entropy, Shannon entropy, collision entropy, and min-entropy. The Rényi entropy is named after Alfréd Rényi, who looked for the most general way to quantify information while preserving additivity for independent events. In the context of fractal dimension estimation, the Rényi entropy forms the basis of the concept of generalized dimensions.
In statistics and information theory, a maximum entropy probability distribution has entropy that is at least as great as that of all other members of a specified class of probability distributions. According to the principle of maximum entropy, if nothing is known about a distribution except that it belongs to a certain class, then the distribution with the largest entropy should be chosen as the least-informative default. The motivation is twofold: first, maximizing entropy minimizes the amount of prior information built into the distribution; second, many physical systems tend to move towards maximal entropy configurations over time.
In information theory, Gibbs' inequality is a statement about the information entropy of a discrete probability distribution. Several other bounds on the entropy of probability distributions are derived from Gibbs' inequality, including Fano's inequality. It was first presented by J. Willard Gibbs in the 19th century.
The principle of detailed balance can be used in kinetic systems which are decomposed into elementary processes. It states that at equilibrium, each elementary process is in equilibrium with its reverse process.
The Theil index is a statistic primarily used to measure economic inequality and other economic phenomena, though it has also been used to measure racial segregation.
In thermodynamics, the entropy of mixing is the increase in the total entropy when several initially separate systems of different composition, each in a thermodynamic state of internal equilibrium, are mixed without chemical reaction by the thermodynamic operation of removal of impermeable partition(s) between them, followed by a time for establishment of a new thermodynamic state of internal equilibrium in the new unpartitioned closed system.
Differential entropy is a concept in information theory that began as an attempt by Claude Shannon to extend the idea of (Shannon) entropy, a measure of average (surprisal) of a random variable, to continuous probability distributions. Unfortunately, Shannon did not derive this formula, and rather just assumed it was the correct continuous analogue of discrete entropy, but it is not. The actual continuous version of discrete entropy is the limiting density of discrete points (LDDP). Differential entropy is commonly encountered in the literature, but it is a limiting case of the LDDP, and one that loses its fundamental association with discrete entropy.
In ecology, alpha diversity (α-diversity) is the mean species diversity in a site at a local scale. The term was introduced by R. H. Whittaker together with the terms beta diversity (β-diversity) and gamma diversity (γ-diversity). Whittaker's idea was that the total species diversity in a landscape is determined by two different things, the mean species diversity in sites at a more local scale and the differentiation among those sites.
The concept entropy was first developed by German physicist Rudolf Clausius in the mid-nineteenth century as a thermodynamic property that predicts that certain spontaneous processes are irreversible or impossible. In statistical mechanics, entropy is formulated as a statistical property using probability theory. The statistical entropy perspective was introduced in 1870 by Austrian physicist Ludwig Boltzmann, who established a new field of physics that provided the descriptive linkage between the macroscopic observation of nature and the microscopic view based on the rigorous treatment of large ensembles of microstates that constitute thermodynamic systems.
In ecology, gamma diversity (γ-diversity) is the total species diversity in a landscape. The term was introduced by R. H. Whittaker together with the terms alpha diversity (α-diversity) and beta diversity (β-diversity). Whittaker's idea was that the total species diversity in a landscape (γ) is determined by two different things, the mean species diversity in sites at a more local scale (α) and the differentiation among those sites (β). According to this reasoning, alpha diversity and beta diversity constitute independent components of gamma diversity:
An index of qualitative variation (IQV) is a measure of statistical dispersion in nominal distributions. There are a variety of these, but they have been relatively little-studied in the statistics literature. The simplest is the variation ratio, while more complex indices include the information entropy.
In mathematics, a function defined on a region of the complex plane is said to be of bounded type if it is equal to the ratio of two analytic functions bounded in that region. But more generally, a function is of bounded type in a region if and only if is analytic on and has a harmonic majorant on where . Being the ratio of two bounded analytic functions is a sufficient condition for a function to be of bounded type, and if is simply connected the condition is also necessary.
The Gibbs rotational ensemble represents the possible states of a mechanical system in thermal and rotational equilibrium at temperature and angular velocity . The Jaynes procedure can be used to obtain this ensemble. An ensemble is the set of microstates corresponding to a given macrostate.
↑ Chao, Anne; Chiu, Chun-Huo; Jost, Lou (2016), "Phylogenetic Diversity Measures and Their Decomposition: A Framework Based on Hill Numbers", Biodiversity Conservation and Phylogenetic Systematics, Topics in Biodiversity and Conservation, Springer International Publishing, vol.14, pp.141–172, doi:10.1007/978-3-319-22461-9_8, ISBN9783319224602
↑ Spellerberg, Ian F., and Peter J. Fedor. (2003) A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’Index. Global Ecology and Biogeography 12.3, 177-179.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.