
Butterflies (Rhopalocera) are insects that have large, often brightly coloured wings, and a conspicuous, fluttering flight. The group comprises the superfamilies Hedyloidea and Papilionoidea. The oldest butterfly fossils have been dated to the Paleocene, about 56 million years ago, though they may have originated earlier.

In evolutionary biology, mimicry is an evolved resemblance between an organism and another object, often an organism of another species. Mimicry may evolve between different species, or between individuals of the same species. Often, mimicry functions to protect a species from predators, making it an anti-predator adaptation. Mimicry evolves if a receiver perceives the similarity between a mimic and a model and as a result changes its behaviour in a way that provides a selective advantage to the mimic. The resemblances that evolve in mimicry can be visual, acoustic, chemical, tactile, or electric, or combinations of these sensory modalities. Mimicry may be to the advantage of both organisms that share a resemblance, in which case it is a form of mutualism; or mimicry can be to the detriment of one, making it parasitic or competitive. The evolutionary convergence between groups is driven by the selective action of a signal-receiver or dupe. Birds, for example, use sight to identify palatable insects and butterflies, whilst avoiding the noxious ones. Over time, palatable insects may evolve to resemble noxious ones, making them mimics and the noxious ones models. In the case of mutualism, sometimes both groups are referred to as "co-mimics". It is often thought that models must be more abundant than mimics, but this is not so. Mimicry may involve numerous species; many harmless species such as hoverflies are Batesian mimics of strongly defended species such as wasps, while many such well-defended species form Müllerian mimicry rings, all resembling each other. Mimicry between prey species and their predators often involves three or more species.

The Gulf fritillary or passion butterfly is a bright orange butterfly in the subfamily Heliconiinae of the family Nymphalidae. That subfamily was formerly set apart as a separate family, the Heliconiidae. The Heliconiinae are "longwing butterflies", which have long, narrow wings compared to other butterflies.
Batesian mimicry is a form of mimicry where a harmless species has evolved to imitate the warning signals of a harmful species directed at a predator of them both. It is named after the English naturalist Henry Walter Bates, after his work on butterflies in the rainforests of Brazil.

The viceroy is a North American butterfly. It was long thought to be a Batesian mimic of the monarch butterfly, but since the viceroy is also distasteful to predators, it is now considered a Müllerian mimic instead.

Heliconius charithonia, the zebra longwing or zebra heliconian, is a species of butterfly belonging to the subfamily Heliconiinae of the family Nymphalidae. It was first described by Carl Linnaeus in his 1767 12th edition of Systema Naturae. The boldly striped black and white wing pattern is aposematic, warning off predators.

Dryas iulia, commonly called the Julia butterfly, Julia heliconian, the flame, or flambeau, is a species of brush-footed butterfly. The sole representative of its genus Dryas, it is native from Brazil to southern Texas and Florida, and in summer can sometimes be found as far north as eastern Nebraska. Over 15 subspecies have been described.

Müllerian mimicry is a natural phenomenon in which two or more well-defended species, often foul-tasting and sharing common predators, have come to mimic each other's honest warning signals, to their mutual benefit. The benefit to Müllerian mimics is that predators only need one unpleasant encounter with one member of a set of Müllerian mimics, and thereafter avoid all similar coloration, whether or not it belongs to the same species as the initial encounter. It is named after the German naturalist Fritz Müller, who first proposed the concept in 1878, supporting his theory with the first mathematical model of frequency-dependent selection, one of the first such models anywhere in biology.

Danaus chrysippus, also known as the plain tiger, African queen, or African monarch, is a medium-sized butterfly widespread in Asia, Australia and Africa. It belongs to the Danainae subfamily of the brush-footed butterfly family Nymphalidae. Danainae primarily consume plants in the genus Asclepias, more commonly called milkweed. Milkweed contains toxic compounds, cardenolides, which are often consumed and stored by many butterflies. Because of their emetic properties, the plain tiger is unpalatable to most predators. As a result, its coloration is widely mimicked by other species of butterflies. The plain tiger inhabits a wide variety of habitats, although it is less likely to thrive in jungle-like conditions and is most often found in drier, wide-open areas.

The queen butterfly is a North and South American butterfly in the family Nymphalidae with a wingspan of 80–85 mm. It is orange or brown with black wing borders and small white forewing spots on its dorsal wing surface, and reddish ventral wing surface fairly similar to the dorsal surface. The ventral hindwings have black veins and small white spots in a black border. The male has a black androconial scent patch on its dorsal hindwings. It can be found in meadows, fields, marshes, deserts, and at the edges of forests.

Heliconius cydno, the cydno longwing, is a nymphalid butterfly that ranges from Mexico to northern South America. It is typically found in the forest understory and deposits its eggs on a variety of plants of the genus Passiflora. It is a member of the Heliconiinae subfamily of Central and South America, and it is the only heliconiine that can be considered oligophagous. H. cydno is also characterized by hybridization and Müllerian mimicry. Wing coloration plays a key role in mate choice and has further implications in regards to sympatric speciation. Macrolide scent gland extracts and wing-clicking behavior further characterize this species.
Heliconius comprises a colorful and widespread genus of brush-footed butterflies commonly known as the longwings or heliconians. This genus is distributed throughout the tropical and subtropical regions of the New World, from South America as far north as the southern United States. The larvae of these butterflies eat passion flower vines (Passifloraceae). Adults exhibit bright wing color patterns which signal their distastefulness to potential predators.

Heliconius melpomene, the postman butterfly, common postman or simply postman, is a brightly colored butterfly found throughout Central and South America. It was first described by Carl Linnaeus in his 1758 10th edition of Systema Naturae. Its coloration coevolved with a sister species H. erato as a warning to predators of its inedibility; this is an example of Müllerian mimicry. H. melpomene was one of the first butterfly species observed to forage for pollen, a behavior that is common in other groups but rare in butterflies. Because of the recent rapid evolutionary radiation of the genus Heliconius and overlapping of its habitat with other related species, H. melpomene has been the subject of extensive study on speciation and hybridization. These hybrids tend to have low fitness as they look different from the original species and no longer exhibit Müllerian mimicry.

Heliconius ismenius, the Ismenius tiger or tiger heliconian, is a butterfly of the family Nymphalidae found in Central America and northern South America. They are abundant as far south as Ecuador and Venezuela and as far north as southern Mexico, Guatemala and Belize. H. ismenius are more commonly called the tiger-striped long wing butterfly. H. ismenius's nickname is derived from its long wing structure as well as the beautiful burnt orange and black stripes. Pierre André Latreille, a French zoologist, described Heliconius ismenius in 1817. H. ismenius resembles a number of other butterflies, an example of Müllerian mimicry.

Heliconius heurippa is a butterfly of the genus Heliconius that is believed by some scientists to be a separate species from—but a hybrid of—the species Heliconius cydno and Heliconius melpomene, making H. heurippa an example of hybrid speciation.

Chemical mimicry is a type of biological mimicry involving the use of chemicals to dupe an operator.

Heliconius numata, the Numata longwing, is a brush-footed butterfly species belonging to the family Nymphalidae, subfamily Heliconiinae.
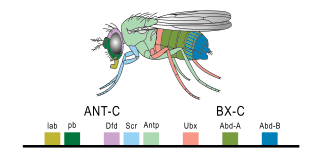
The evo-devo gene toolkit is the small subset of genes in an organism's genome whose products control the organism's embryonic development. Toolkit genes are central to the synthesis of molecular genetics, palaeontology, evolution and developmental biology in the science of evolutionary developmental biology (evo-devo). Many of them are ancient and highly conserved among animal phyla.

Heliconius eleuchia, the white-edged longwing, is a species of Heliconius butterfly described by William Chapman Hewitson in 1853.
Heliconius erato petiverana is a subspecies or geographical race of the widespread neotropical nymphalid butterfly, Heliconius erato. It is characterized by a red forewing patch and a yellow transverse stripe on the hindwing, on an otherwise black or dark brown background.The type locality is "Mexico", and the form occurs from southern Texas to central Panama. Some lepidopterists distinguish the named forms H. erato cruentus and H. erato demophoon as separate races, based on the size of the red FW patch and the thickness of the yellow HW stripe.












