Related Research Articles

Symbiosis is any type of a close and long-term biological interaction between two biological organisms of different species, termed symbionts, be it mutualistic, commensalistic, or parasitic. In 1879, Heinrich Anton de Bary defined it as "the living together of unlike organisms". The term is sometimes used in the more restricted sense of a mutually beneficial interaction in which both symbionts contribute to each other's support.
Horizontal transmission is the transmission of organisms between biotic and/or abiotic members of an ecosystem that are not in a parent-progeny relationship. This concept has been generalized to include transmissions of infectious agents, symbionts, and cultural traits between humans.

Microbiota are the range of microorganisms that may be commensal, mutualistic, or pathogenic found in and on all multicellular organisms, including plants. Microbiota include bacteria, archaea, protists, fungi, and viruses, and have been found to be crucial for immunologic, hormonal, and metabolic homeostasis of their host.
For the American folk-rock singer-songwriter, see Nancy Moran.
The hologenome theory of evolution recasts the individual animal or plant as a community or a "holobiont" – the host plus all of its symbiotic microbes. Consequently, the collective genomes of the holobiont form a "hologenome". Holobionts and hologenomes are structural entities that replace misnomers in the context of host-microbiota symbioses such as superorganism, organ, and metagenome. Variation in the hologenome may encode phenotypic plasticity of the holobiont and can be subject to evolutionary changes caused by selection and drift, if portions of the hologenome are transmitted between generations with reasonable fidelity. One of the important outcomes of recasting the individual as a holobiont subject to evolutionary forces is that genetic variation in the hologenome can be brought about by changes in the host genome and also by changes in the microbiome, including new acquisitions of microbes, horizontal gene transfers, and changes in microbial abundance within hosts. Although there is a rich literature on binary host–microbe symbioses, the hologenome concept distinguishes itself by including the vast symbiotic complexity inherent in many multicellular hosts. For recent literature on holobionts and hologenomes published in an open access platform, see the following reference.

The root microbiome is the dynamic community of microorganisms associated with plant roots. Because they are rich in a variety of carbon compounds, plant roots provide unique environments for a diverse assemblage of soil microorganisms, including bacteria, fungi, and archaea. The microbial communities inside the root and in the rhizosphere are distinct from each other, and from the microbial communities of bulk soil, although there is some overlap in species composition.
The phycosphere is a microscale mucus region that is rich in organic matter surrounding a phytoplankton cell. This area is high in nutrients due to extracellular waste from the phytoplankton cell and it has been suggested that bacteria inhabit this area to feed on these nutrients. This high nutrient environment creates a microbiome and a diverse food web for microbes such as bacteria and protists. It has also been suggested that the bacterial assemblages within the phycosphere are species-specific and can vary depending on different environmental factors.
A microbiome is the community of microorganisms that can usually be found living together in any given habitat. It was defined more precisely in 1988 by Whipps et al. as "a characteristic microbial community occupying a reasonably well-defined habitat which has distinct physio-chemical properties. The term thus not only refers to the microorganisms involved but also encompasses their theatre of activity". In 2020, an international panel of experts published the outcome of their discussions on the definition of the microbiome. They proposed a definition of the microbiome based on a revival of the "compact, clear, and comprehensive description of the term" as originally provided by Whipps et al., but supplemented with two explanatory paragraphs. The first explanatory paragraph pronounces the dynamic character of the microbiome, and the second explanatory paragraph clearly separates the term microbiota from the term microbiome.
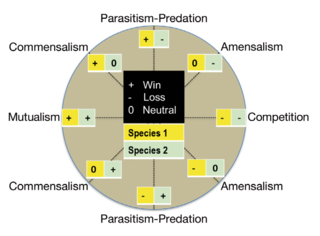
Microbial symbiosis in marine animals was not discovered until 1981. In the time following, symbiotic relationships between marine invertebrates and chemoautotrophic bacteria have been found in a variety of ecosystems, ranging from shallow coastal waters to deep-sea hydrothermal vents. Symbiosis is a way for marine organisms to find creative ways to survive in a very dynamic environment. They are different in relation to how dependent the organisms are on each other or how they are associated. It is also considered a selective force behind evolution in some scientific aspects. The symbiotic relationships of organisms has the ability to change behavior, morphology and metabolic pathways. With increased recognition and research, new terminology also arises, such as holobiont, which the relationship between a host and its symbionts as one grouping. Many scientists will look at the hologenome, which is the combined genetic information of the host and its symbionts. These terms are more commonly used to describe microbial symbionts.

Eugene Rosenberg is a microbiologist at the Faculty of Life Sciences at Tel Aviv University, an expert in the field of applied environmental microbiology, in particular his work on Myxobacteria, microorganisms to combat pollution (bioremediation), and the Hologenome theory of evolution.
A holobiont is an assemblage of a host and the many other species living in or around it, which together form a discrete ecological unit through symbiosis, though there is controversy over this discreteness. The components of a holobiont are individual species or bionts, while the combined genome of all bionts is the hologenome. The holobiont concept was initially introduced by the German theoretical biologist Adolf Meyer-Abich in 1943, and then apparently independently by Dr. Lynn Margulis in her 1991 book Symbiosis as a Source of Evolutionary Innovation. The concept has evolved since the original formulations. Holobionts include the host, virome, microbiome, and any other organisms which contribute in some way to the functioning of the whole. Well-studied holobionts include reef-building corals and humans.
Social selection is a term used with varying meanings in biology.
Hologenomics is the omics study of hologenomes. A hologenome is the whole set of genomes of a holobiont, an organism together with all co-habitating microbes, other life forms, and viruses. While the term hologenome originated from the hologenome theory of evolution, which postulates that natural selection occurs on the holobiont level, hologenomics uses an integrative framework to investigate interactions between the host and its associated species. Examples include gut microbe or viral genomes linked to human or animal genomes for host-microbe interaction research. Hologenomics approaches have also been used to explain genetic diversity in the microbial communities of marine sponges.
Vertical transmission of symbionts is the transfer of a microbial symbiont from the parent directly to the offspring. Many metazoan species carry symbiotic bacteria which play a mutualistic, commensal, or parasitic role. A symbiont is acquired by a host via horizontal, vertical, or mixed transmission.

All animals on Earth form associations with microorganisms, including protists, bacteria, archaea, fungi, and viruses. In the ocean, animal–microbial relationships were historically explored in single host–symbiont systems. However, new explorations into the diversity of marine microorganisms associating with diverse marine animal hosts is moving the field into studies that address interactions between the animal host and a more multi-member microbiome. The potential for microbiomes to influence the health, physiology, behavior, and ecology of marine animals could alter current understandings of how marine animals adapt to change, and especially the growing climate-related and anthropogenic-induced changes already impacting the ocean environment.
The plant microbiome, also known as the phytomicrobiome, plays roles in plant health and productivity and has received significant attention in recent years. The microbiome has been defined as "a characteristic microbial community occupying a reasonably well-defined habitat which has distinct physio-chemical properties. The term thus not only refers to the microorganisms involved but also encompasses their theatre of activity".
Since the colonization of land by ancestral plant lineages 450 million years ago, plants and their associated microbes have been interacting with each other, forming an assemblage of species that is often referred to as a holobiont. Selective pressure acting on holobiont components has likely shaped plant-associated microbial communities and selected for host-adapted microorganisms that impact plant fitness. However, the high microbial densities detected on plant tissues, together with the fast generation time of microbes and their more ancient origin compared to their host, suggest that microbe-microbe interactions are also important selective forces sculpting complex microbial assemblages in the phyllosphere, rhizosphere, and plant endosphere compartments.
The holobiont concept is a renewed paradigm in biology that can help to describe and understand complex systems, like the host-microbe interactions that play crucial roles in marine ecosystems. However, there is still little understanding of the mechanisms that govern these relationships, the evolutionary processes that shape them and their ecological consequences. The holobiont concept posits that a host and its associated microbiota with which it interacts, form a holobiont, and have to be studied together as a coherent biological and functional unit to understand its biology, ecology, and evolution.
Catalina Cuellar-Gempeler is a Colombian microbial ecologist and marine microbiologist, currently teaching and doing research at Cal Poly Humboldt. Her research focuses mainly on understanding microbial meta-community, eco-evolutionary dynamics, and ecosystem dynamics. The Catalina Cuellar-Gempeler lab is currently focused on studying the interactions between hosts and their microbial communities. The lab's main emphasis is on the microbes used in digestion in the Californian and Eastern carnivorous pitcher plants. In March 2021, Cuellar-Gempeler was awarded an Early Career grant of $1 million by the National Science Foundation.
Megan Frederickson is a Canadian evolutionary biologist who is a professor of Ecology and Evolutionary Biology at the University of Toronto. Her research considers the evolution of cooperation and the ecological genetics of mutualism.
References
- ↑ Yoon, Carol Kaesuk (October 17, 2000). "Scientist at Work: Joan Roughgarden; A Theorist With Personal Experience Of the Divide Between the Sexes". The New York Times. Retrieved March 7, 2014.
- ↑ "Curriculum Vitae: Joan Roughgarden". Stanford University. Retrieved March 7, 2014.
- ↑ Pacala, Stephen W.; Roughgarden, Jonathan (1985). "Population Experiments with the Anolis Lizards of St. Maarten and St. Eustatius". Ecology. 66 (1): 129–141. doi:10.2307/1941313. ISSN 1939-9170. JSTOR 1941313.
- ↑ Post, David M.; Palkovacs, Eric P. (June 12, 2009). "Eco-evolutionary feedbacks in community and ecosystem ecology: interactions between the ecological theatre and the evolutionary play". Philosophical Transactions of the Royal Society B: Biological Sciences. 364 (1523): 1629–1640. doi:10.1098/rstb.2009.0012. ISSN 0962-8436. PMC 2690506 . PMID 19414476.
- ↑ Paine, Robert T.; Vadas, Robert L. (1969). "The Effects of Grazing by Sea Urchins, Strongylocentrotus spp., on Benthic Algal Populations". Limnology and Oceanography. 14 (5): 710–719. Bibcode:1969LimOc..14..710P. doi: 10.4319/lo.1969.14.5.0710 .
- 1 2 Connell, Joseph H. (1961). "The Influence of Interspecific Competition and Other Factors on the Distribution of the Barnacle Chthamalus Stellatus". Ecology. 42 (4): 710–723. doi:10.2307/1933500. ISSN 0012-9658. JSTOR 1933500.
- ↑ Connell, Joseph H. (1961). "Effects of Competition, Predation by Thais lapillus, and Other Factors on Natural Populations of the Barnacle Balanus balanoides" . Ecological Monographs. 31 (1): 61–104. doi:10.2307/1950746. ISSN 0012-9615. JSTOR 1950746.
- 1 2 Gaines, S.; Roughgarden, J. (June 1, 1985). "Larval settlement rate: A leading determinant of structure in an ecological community of the marine intertidal zone". Proceedings of the National Academy of Sciences. 82 (11): 3707–3711. Bibcode:1985PNAS...82.3707G. doi: 10.1073/pnas.82.11.3707 . ISSN 0027-8424. PMC 397856 . PMID 16593571.
- ↑ Connolly, Sean R.; Roughgarden, Jonathan (April 1, 1998). "A Latitudinal Gradient in Northeast Pacific Intertidal Community Structure: Evidence for an Oceanographically Based Synthesis of Marine Community Theory". The American Naturalist. 151 (4): 311–326. doi:10.1086/286121. ISSN 0003-0147. PMID 18811323. S2CID 23831003.
- ↑ Connolly, Sean R.; Roughgarden, Joan (1999). "Theory of Marine Communities: Competition, Predation, and Recruitment-Dependent Interaction Strength". Ecological Monographs. 69 (3): 277–296. doi:10.1890/0012-9615(1999)069[0277:tomccp]2.0.co;2. ISSN 0012-9615.
- ↑ Caley, M. J.; Carr, M. H.; Hixon, M. A.; Hughes, T. P.; Jones, G. P.; Menge, B. A. (1996). "Recruitment and the Local Dynamics of Open Marine Populations". Annual Review of Ecology and Systematics. 27 (1): 477–500. doi:10.1146/annurev.ecolsys.27.1.477. ISSN 0066-4162. Archived from the original on January 31, 2022. Retrieved January 31, 2022.
- ↑ Roughgarden, Joan (2013). Evolution's rainbow : diversity, gender, and sexuality in nature and people (Tenth anniversary ed.). Berkeley. ISBN 978-0-520-95797-8. OCLC 861528157.
{{cite book}}: CS1 maint: location missing publisher (link) - ↑ Moser, Bob. "On the Originality of Species". Stanford Magazine. No. May/June 2004. Stanford University . Retrieved June 13, 2021.
- ↑ Jonah Lehrer. "The Effeminate Sheep". Seed (June/July 2006). Reprinted in Richard Preston, ed. (2007). The Best American Science and Nature Writing. New York: Houghton Mifflin. ISBN 978-0-618-72231-0.
- 1 2 "Q&A with Joan Roughgarden on the Problems with the Theory of Sexual Selection". MIT Press . February 11, 2019. Archived from the original on June 13, 2021. Retrieved June 13, 2021.
- 1 2 3 Roughgarden, Joan; Oishi, Meeko; Akçay, Erol (February 17, 2006). "Reproductive Social Behavior: Cooperative Games to Replace Sexual Selection". Science. 311 (5763): 965–969. Bibcode:2006Sci...311..965R. doi:10.1126/science.1110105. ISSN 0036-8075. PMID 16484485. S2CID 32364112.
- 1 2 3 4 Atkinson, Nick (May 5, 2006). "Sexual selection alternative slammed". The Scientist . Archived from the original on September 29, 2007. Retrieved May 15, 2007.
- 1 2 3 Dall, S. R. X.; McNamara, J. M.; Wedell, N.; Hosken, D. J. (May 5, 2006). "Debating Sexual Selection and Mating Strategies" (PDF). Science. 312 (5774): 689b–697b. doi:10.1126/science.312.5774.689b. PMID 16675684. S2CID 10887773 . Retrieved June 14, 2021.
- 1 2 Clutton-Brock, Tim (December 21, 2007). "Sexual Selection in Males and Females". Science. 318 (5858): 1882–1885. Bibcode:2007Sci...318.1882C. doi:10.1126/science.1133311. ISSN 0036-8075. PMID 18096798. S2CID 6883765.
- ↑ Roughgarden, Joan (2009). The genial gene : deconstructing Darwinian selfishness. Berkeley: University of California Press. ISBN 978-0-520-25826-6. OCLC 264039560.
- ↑ Dawkins, Richard. The selfish gene. OCLC 1238007082.
- ↑ Folse HJ, 3rd; Roughgarden, J (2010). "What is an individual organism? A multilevel selection perspective". The Quarterly Review of Biology. 85 (4): 447–72. doi:10.1086/656905. PMID 21243964. S2CID 19816447.
{{cite journal}}: CS1 maint: numeric names: authors list (link) - ↑ Roughgarden, J. (2012). "The social selection alternative to sexual selection". Philosophical Transactions of the Royal Society B: Biological Sciences. 367 (1600): 2294–2303. doi:10.1098/rstb.2011.0282. PMC 3391423 . PMID 22777017.
- ↑ Allen, Arthur (May 1, 2014). "Sexual Selection's Mystique Lingers". BioScience. 64 (5): 375–380. doi: 10.1093/biosci/biu047 . ISSN 0006-3568.
- 1 2 Roughgarden, Joan; Adkins-Regan, Elizabeth; Akçay, Erol; Crawford, Jeremy Chase; Gadagkar, Raghavendra; Griffith, Simon C.; Hinde, Camilla A.; Hoquet, Thierry; O’Connor, Cailin; Prokop, Zofia M.; Prum, Richard O. (January 10, 2015), Sexual selection studies: a NESCent catalyst meeting, doi: 10.7287/peerj.preprints.680v3 , S2CID 14039177
- ↑ "First Congregational Church of Berkeley: Event Details". Archived from the original on October 8, 2007. Retrieved September 19, 2006.
- ↑ Roughgarden, Joan. Evolution and Christian Faith: Reflections of an Evolutionary Biologist. Washington D.C.: Island Press, 2006.
- ↑ "TSN: Beyond Belief: Science, Reason, Religion & Survival". thesciencenetwork.org. Retrieved September 27, 2016.
- 1 2 3 Roughgarden, Joan; Gilbert, Scott F.; Rosenberg, Eugene; Zilber-Rosenberg, Ilana; Lloyd, Elisabeth A. (2018). "Holobionts as Units of Selection and a Model of Their Population Dynamics and Evolution". Biological Theory. 13 (1): 44–65. doi: 10.1007/s13752-017-0287-1 . ISSN 1555-5542. S2CID 256282704.
- ↑ Bordenstein, Seth R.; Theis, Kevin R. (August 18, 2015). "Host Biology in Light of the Microbiome: Ten Principles of Holobionts and Hologenomes". PLOS Biology. 13 (8): e1002226. doi: 10.1371/journal.pbio.1002226 . ISSN 1545-7885. PMC 4540581 . PMID 26284777.
- ↑ Rosenberg, Eugene; Zilber-Rosenberg, Ilana (May 4, 2016). Collier, R. John (ed.). "Microbes Drive Evolution of Animals and Plants: the Hologenome Concept". mBio. 7 (2): e01395. doi:10.1128/mBio.01395-15. ISSN 2161-2129. PMC 4817260 . PMID 27034283.
- ↑ Gilbert, Scott F.; Sapp, Jan; Tauber, Alfred I. (2012). "A Symbiotic View of Life: We Have Never Been Individuals". The Quarterly Review of Biology. 87 (4): 325–341. doi:10.1086/668166. ISSN 0033-5770. PMID 23397797. S2CID 14279096.
- ↑ Moran, Nancy A.; Sloan, Daniel B. (December 4, 2015). "The Hologenome Concept: Helpful or Hollow?". PLOS Biology. 13 (12): e1002311. doi: 10.1371/journal.pbio.1002311 . ISSN 1545-7885. PMC 4670207 . PMID 26636661.
- ↑ Yoshida, Erina; Sakurama, Haruko; Kiyohara, Masashi; Nakajima, Masahiro; Kitaoka, Motomitsu; Ashida, Hisashi; Hirose, Junko; Katayama, Takane; Yamamoto, Kenji; Kumagai, Hidehiko (September 16, 2011). "Bifidobacterium longum subsp. infantis uses two different β-galactosidases for selectively degrading type-1 and type-2 human milk oligosaccharides". Glycobiology. 22 (3): 361–368. doi: 10.1093/glycob/cwr116 . ISSN 1460-2423. PMID 21926104.
- ↑ Dupressoir, A.; Lavialle, C.; Heidmann, T. (2012). "From ancestral infectious retroviruses to bona fide cellular genes: Role of the captured syncytins in placentation". Placenta. 33 (9): 663–671. doi:10.1016/j.placenta.2012.05.005. ISSN 0143-4004. PMID 22695103.
- 1 2 Roughgarden, Joan (March 2, 2020). "Holobiont Evolution: Mathematical Model with Vertical vs. Horizontal Microbiome Transmission". Philosophy, Theory, and Practice in Biology. 12 (20220112). doi: 10.3998/ptpbio.16039257.0012.002 . hdl:2027/spo.16039257.0012.002. ISSN 2475-3025. S2CID 216329421.
- 1 2 3 Roughgarden, Joan (June 1, 2023). "Holobiont Evolution: Population Theory for the Hologenome". The American Naturalist. 201 (6): 763–778. doi: 10.1086/723782 . ISSN 0003-0147. PMID 37229712. S2CID 220253227.
- ↑ "Lloyd W. Dinkelspiel Awards | Registrar's Office". registrar.stanford.edu. Retrieved April 4, 2019.
- ↑ "Science Matters". California Science Center. Archived from the original on July 13, 2010.