
Gestation is the period of development during the carrying of an embryo, and later fetus, inside viviparous animals. It is typical for mammals, but also occurs for some non-mammals. Mammals during pregnancy can have one or more gestations at the same time, for example in a multiple birth.

The uterus or womb is the organ in the reproductive system of most female mammals, including humans, that accommodates the embryonic and fetal development of one or more embryos until birth. The uterus is a hormone-responsive sex organ that contains glands in its lining that secrete uterine milk for embryonic nourishment.

The placenta is a temporary embryonic and later fetal organ that begins developing from the blastocyst shortly after implantation. It plays critical roles in facilitating nutrient, gas and waste exchange between the physically separate maternal and fetal circulations, and is an important endocrine organ, producing hormones that regulate both maternal and fetal physiology during pregnancy. The placenta connects to the fetus via the umbilical cord, and on the opposite aspect to the maternal uterus in a species-dependent manner. In humans, a thin layer of maternal decidual (endometrial) tissue comes away with the placenta when it is expelled from the uterus following birth. Placentas are a defining characteristic of placental mammals, but are also found in marsupials and some non-mammals with varying levels of development.
The oviduct in mammals is the passageway from an ovary. In human females, this is more usually known as the fallopian tube or uterine tube. The eggs travel along the oviduct. These eggs will either be fertilized by spermatozoa to become a zygote, or will degenerate in the body. Normally, these are paired structures, but in birds and some cartilaginous fishes, one or the other side fails to develop, and only one functional oviduct can be found.

The amnion is a membrane that closely covers the human and various other embryos when first formed. It fills with amniotic fluid, which causes the amnion to expand and become the amniotic sac that provides a protective environment for the developing embryo. The amnion, along with the chorion, the yolk sac and the allantois protect the embryo. In birds, reptiles and monotremes, the protective sac is enclosed in a shell. In marsupials and placental mammals, it is enclosed in a uterus.

In animals, viviparity is development of the embryo inside the body of the mother, with the maternal circulation providing for the metabolic needs of the embryo's development, until the mother gives birth to a fully or partially developed juvenile that is at least metabolically independent. This is opposed to oviparity, where the embryos develop independently outside the mother in eggs until they are developed enough to break out as hatchlings; and ovoviviparity, where the embryos are developed in eggs that remain carried inside the mother's body until the hatchlings emerge from the mother as juveniles, similar to a live birth.

Gestational hypertension or pregnancy-induced hypertension (PIH) is the development of new hypertension in a pregnant woman after 20 weeks' gestation without the presence of protein in the urine or other signs of pre-eclampsia. Gestational hypertension is defined as having a blood pressure greater than 140/90 on two occasions at least 6 hours apart.

The trophoblast is the outer layer of cells of the blastocyst. Trophoblasts are present four days after fertilization in humans. They provide nutrients to the embryo and develop into a large part of the placenta. They form during the first stage of pregnancy and are the first cells to differentiate from the fertilized egg to become extraembryonic structures that do not directly contribute to the embryo. After blastulation, the trophoblast is contiguous with the ectoderm of the embryo and is referred to as the trophectoderm. After the first differentiation, the cells in the human embryo lose their totipotency because they can no longer form a trophoblast. They become pluripotent stem cells.
Antepartum bleeding, also known as antepartum haemorrhage (APH) or prepartum hemorrhage, is genital bleeding during pregnancy after the 28th week of pregnancy up to delivery.

The decidua is the modified mucosal lining of the uterus that forms every month, in preparation for pregnancy. It is shed off each month when there is no fertilised egg to support. The decidua is under the influence of progesterone. Endometrial cells become highly characteristic. The decidua forms the maternal part of the placenta and remains for the duration of the pregnancy. After birth the decidua is shed together with the placenta.
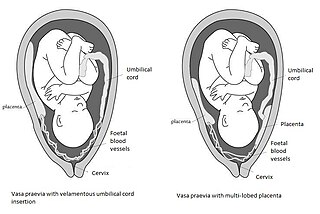
Vasa praevia is a condition in which fetal blood vessels cross or run near the internal opening of the uterus. These vessels are at risk of rupture when the supporting membranes rupture, as they are unsupported by the umbilical cord or placental tissue.

Implantation, also known as nidation, is the stage in the embryonic development of mammals in which the blastocyst hatches, attaches, adheres, and invades into the wall of the female's uterus. Implantation is the first stage of gestation, and, when successful, the female is considered to be pregnant. An implanted embryo is detected by the presence of increased levels of human chorionic gonadotropin (hCG) in a pregnancy test. The implanted embryo will receive oxygen and nutrients in order to grow.
Decidualization is a process that results in significant changes to cells of the endometrium in preparation for, and during, pregnancy. This includes morphological and functional changes to endometrial stromal cells (ESCs), the presence of decidual white blood cells (leukocytes), and vascular changes to maternal arteries. The sum of these changes results in the endometrium changing into a structure called the decidua. In humans, the decidua is shed during childbirth.

Uterine glands or endometrial glands are tubular glands, lined by a simple columnar epithelium, found in the functional layer of the endometrium that lines the uterus. Their appearance varies during the menstrual cycle. During the proliferative phase, uterine glands appear long due to estrogen secretion by the ovaries. During the secretory phase, the uterine glands become very coiled with wide lumens and produce a glycogen-rich secretion known as histotroph or uterine milk. This change corresponds with an increase in blood flow to spiral arteries due to increased progesterone secretion from the corpus luteum. During the pre-menstrual phase, progesterone secretion decreases as the corpus luteum degenerates, which results in decreased blood flow to the spiral arteries. The functional layer of the uterus containing the glands becomes necrotic, and eventually sloughs off during the menstrual phase of the cycle.

Pseudemoia entrecasteauxii, also known commonly as Entrecasteaux's skink, the southern grass skink, the tussock cool-skink, and the tussock skink, is a species of lizard in the family Scincidae. The species is endemic to Australia.
Immune tolerance in pregnancy or maternal immune tolerance is the immune tolerance shown towards the fetus and placenta during pregnancy. This tolerance counters the immune response that would normally result in the rejection of something foreign in the body, as can happen in cases of spontaneous abortion. It is studied within the field of reproductive immunology.

The fetal membranes are the four extraembryonic membranes, associated with the developing embryo, and fetus in humans and other mammals. They are the amnion, chorion, allantois, and yolk sac. The amnion and the chorion are the chorioamniotic membranes that make up the amniotic sac which surrounds and protects the embryo. The fetal membranes are four of six accessory organs developed by the conceptus that are not part of the embryo itself, the other two are the placenta, and the umbilical cord.

Placental expulsion occurs when the placenta comes out of the birth canal after childbirth. The period from just after the baby is expelled until just after the placenta is expelled is called the third stage of labor.

Most mammals are viviparous, giving birth to live young. However, the five species of monotreme, the platypuses and the echidnas, lay eggs. The monotremes have a sex determination system different from that of most other mammals. In particular, the sex chromosomes of a platypus are more like those of a chicken than those of a therian mammal.

Pregnancy has been traditionally defined as the period of time eggs are incubated in the body after the egg-sperm union. Although the term often refers to placental mammals, it has also been used in the titles of many international, peer-reviewed, scientific articles on fish, e.g. Consistent with this definition, there are several modes of reproduction in fish, providing different amounts of parental care. In ovoviviparity, there is internal fertilization and the young are born live but there is no placental connection or significant trophic (feeding) interaction; the mother's body maintains gas exchange but the unborn young are nourished by egg yolk. There are two types of viviparity in fish. In histotrophic viviparity, the zygotes develop in the female's oviducts, but she provides no direct nutrition; the embryos survive by eating her eggs or their unborn siblings. In hemotrophic viviparity, the zygotes are retained within the female and are provided with nutrients by her, often through some form of placenta.





