
A diatom is any member of a large group comprising several genera of algae, specifically microalgae, found in the oceans, waterways and soils of the world. Living diatoms make up a significant portion of the Earth's biomass: they generate about 20 to 50 percent of the oxygen produced on the planet each year, take in over 6.7 billion tonnes of silicon each year from the waters in which they live, and constitute nearly half of the organic material found in the oceans. The shells of dead diatoms can reach as much as a half-mile deep on the ocean floor, and the entire Amazon basin is fertilized annually by 27 million tons of diatom shell dust transported by transatlantic winds from the African Sahara, much of it from the Bodélé Depression, which was once made up of a system of fresh-water lakes.

An algal bloom or algae bloom is a rapid increase or accumulation in the population of algae in freshwater or marine water systems. It is often recognized by the discoloration in the water from the algae's pigments. The term algae encompasses many types of aquatic photosynthetic organisms, both macroscopic multicellular organisms like seaweed and microscopic unicellular organisms like cyanobacteria. Algal bloom commonly refers to the rapid growth of microscopic unicellular algae, not macroscopic algae. An example of a macroscopic algal bloom is a kelp forest.
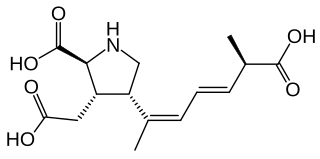
Domoic acid (DA) is a kainic acid-type neurotoxin that causes amnesic shellfish poisoning (ASP). It is produced by algae and accumulates in shellfish, sardines, and anchovies. When sea lions, otters, cetaceans, humans, and other predators eat contaminated animals, poisoning may result. Exposure to this compound affects the brain, causing seizures, and possibly death.

Phaeodactylum tricornutum is a diatom. It is the only species in the genus Phaeodactylum. Unlike other diatoms, P. tricornutum can exist in different morphotypes and changes in cell shape can be stimulated by environmental conditions. This feature can be used to explore the molecular basis of cell shape control and morphogenesis. Unlike most diatoms, P. tricornutum can grow in the absence of silicon and can survive without making silicified frustules. This provides opportunities for experimental exploration of silicon-based nanofabrication in diatoms.
Amnesic shellfish poisoning (ASP) is an illness caused by consumption of shellfish that contain the marine biotoxin called domoic acid. In mammals, including humans, domoic acid acts as a neurotoxin, causing permanent short-term memory loss, brain damage, and death in severe cases.

Øjvind Moestrup is a Danish aquatic botanist, working particularly with the classification of algae. He worked at the Botanical Institute at the University of Copenhagen and is a professor emeritus in the Department of Biology there. He published over 100 scientific papers.

Thalassiosira pseudonana is a species of marine centric diatoms. It was chosen as the first eukaryotic marine phytoplankton for whole genome sequencing. T. pseudonana was selected for this study because it is a model for diatom physiology studies, belongs to a genus widely distributed throughout the world's oceans, and has a relatively small genome at 34 mega base pairs. Scientists are researching on diatom light absorption, using the marine diatom of Thalassiosira. The diatom requires a high enough concentration of CO2 in order to utilize C4 metabolism (Clement et al. 2015).

The genus Ceratium is restricted to a small number of freshwater dinoflagellate species. Previously the genus contained also a large number of marine dinoflagellate species. However, these marine species have now been assigned to a new genus called Tripos. Ceratium dinoflagellates are characterized by their armored plates, two flagella, and horns. They are found worldwide and are of concern due to their blooms.

Chaetoceros is a genus of diatoms in the family Chaetocerotaceae, first described by the German naturalist C. G. Ehrenberg in 1844. Species of this genus are mostly found in marine habitats, but a few species exist in freshwater. It is arguably the common and most diverse genus of marine planktonic diatoms, with over 200 accepted species. It is the type genus of its family.

A harmful algal bloom (HAB), or excessive algae growth, is an algal bloom that causes negative impacts to other organisms by production of natural algae-produced toxins, mechanical damage to other organisms, or by other means. HABs are sometimes defined as only those algal blooms that produce toxins, and sometimes as any algal bloom that can result in severely lower oxygen levels in natural waters, killing organisms in marine or fresh waters. Blooms can last from a few days to many months. After the bloom dies, the microbes that decompose the dead algae use up more of the oxygen, generating a "dead zone" which can cause fish die-offs. When these zones cover a large area for an extended period of time, neither fish nor plants are able to survive. Harmful algal blooms in marine environments are often called "red tides".

Nitzschia is a common pennate marine diatom. In the scientific literature, this genus, named after Christian Ludwig Nitzsch, is sometimes referred to incorrectly as Nitzchia, and it has many species described, which all have a similar morphology. In its current circumscription, Nitzschia is paraphyletic.
Grethe Berit Rytter Hasle was a Norwegian planktologist. Among the first female professors of natural science at the University of Oslo, she specialized in the study of phytoplankton.
Phycotoxins are complex allelopathic chemicals produced by eukaryotic and prokaryotic algal secondary metabolic pathways. More simply, these are toxic chemicals synthesized by photosynthetic organisms. These metabolites are not harmful to the producer but may be toxic to either one or many members of the marine food web. This page focuses on phycotoxins produced by marine microalgae; however, freshwater algae and macroalgae are known phycotoxin producers and may exhibit analogous ecological dynamics. In the pelagic marine food web, phytoplankton are subjected to grazing by macro- and micro-zooplankton as well as competition for nutrients with other phytoplankton species. Marine bacteria try to obtain a share of organic carbon by maintaining symbiotic, parasitic, commensal, or predatory interactions with phytoplankton. Other bacteria will degrade dead phytoplankton or consume organic carbon released by viral lysis. The production of toxins is one strategy that phytoplankton use to deal with this broad range of predators, competitors, and parasites. Smetacek suggested that "planktonic evolution is ruled by protection and not competition. The many shapes of plankton reflect defense responses to specific attack systems". Indeed, phytoplankton retain an abundance of mechanical and chemical defense mechanisms including cell walls, spines, chain/colony formation, and toxic chemical production. These morphological and physiological features have been cited as evidence for strong predatory pressure in the marine environment. However, the importance of competition is also demonstrated by the production of phycotoxins that negatively impact other phytoplankton species. Flagellates are the principle producers of phycotoxins; however, there are known toxigenic diatoms, cyanobacteria, prymnesiophytes, and raphidophytes. Because many of these allelochemicals are large and energetically expensive to produce, they are synthesized in small quantities. However, phycotoxins are known to accumulate in other organisms and can reach high concentrations during algal blooms. Additionally, as biologically active metabolites, phycotoxins may produce ecological effects at low concentrations. These effects may be subtle, but have the potential to impact the biogeographic distributions of phytoplankton and bloom dynamics.

Ditylum brightwellii is a species of cosmopolitan marine centric diatoms. It is a unicellular photosynthetic autotroph that has the ability to divide rapidly and contribute to spring phytoplankton blooms.

Chaetoceros elegans is a species of diatom in the family Chaetocerotaceae. According to Li, 2017, the type locality is Dapeng Bay, Guangdong Province, P. R. China.

Eucampia zodiacus is a common marine centric diatom species. It is known to be cosmopolitan except in polar regions. E. zodiacus is a harmful diatom that has become known as the predominant organisms causing the bleaching of aqua-cultured nori seaweed.
Pseudo-nitzschia australis is a pennate diatom found in temperate and sub-tropic marine waters, such as off the coast of California and Argentina. This diatom is a Harmful Micro Algae that produces toxic effects on a variety of organisms through its production of domoic acid, a neurotoxin. Toxic effects have been observed in a variety of predatory organisms such as pelicans, sea lions, and humans. If exposed to a high enough dose, these predators will die as a result, and there is no known antidote. The potential indirect mortality associated with P. australis is of great concern to humans as toxic algae blooms, including blooms of P. australis, continue to increase in frequency and severity over recent years. Blooms of P. australis are believed to result from high concentrations of nitrates and phosphates in stream and river runoff, as well as coastal upwelling, which are also sources of other harmful algae blooms.

Marine protists are defined by their habitat as protists that live in marine environments, that is, in the saltwater of seas or oceans or the brackish water of coastal estuaries. Life originated as marine single-celled prokaryotes and later evolved into more complex eukaryotes. Eukaryotes are the more developed life forms known as plants, animals, fungi and protists. Protists are the eukaryotes that cannot be classified as plants, fungi or animals. They are mostly single-celled and microscopic. The term protist came into use historically as a term of convenience for eukaryotes that cannot be strictly classified as plants, animals or fungi. They are not a part of modern cladistics because they are paraphyletic.
Greta Albrecht Fryxell was a marine scientist known for her work on the biology and taxonomy of diatoms. In 1996, she was elected a fellow of the American Association for the Advancement of Science.
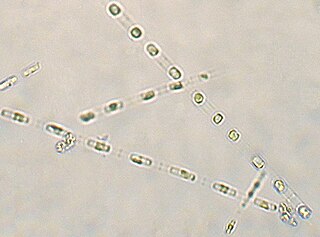
Skeletonema costatum is a cosmopolitan centric diatom that belongs to the genus Skeletonema. It was first described by R. K. Greville, who originally named it Melosira costata, in 1866. It was later renamed by Cleve in 1873 and was more narrowly defined by Zingone et al. and Sarno et al. Skeletonemacostatum is the most well known species of the genus Skeletonema and is often one of the dominant species responsible for red tide events.



