Related Research Articles

The visual cortex of the brain is the area of the cerebral cortex that processes visual information. It is located in the occipital lobe. Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex. The area of the visual cortex that receives the sensory input from the lateral geniculate nucleus is the primary visual cortex, also known as visual area 1 (V1), Brodmann area 17, or the striate cortex. The extrastriate areas consist of visual areas 2, 3, 4, and 5.

The visual system comprises the sensory organ and parts of the central nervous system which gives organisms the sense of sight as well as enabling the formation of several non-image photo response functions. It detects and interprets information from the optical spectrum perceptible to that species to "build a representation" of the surrounding environment. The visual system carries out a number of complex tasks, including the reception of light and the formation of monocular neural representations, colour vision, the neural mechanisms underlying stereopsis and assessment of distances to and between objects, the identification of a particular object of interest, motion perception, the analysis and integration of visual information, pattern recognition, accurate motor coordination under visual guidance, and more. The neuropsychological side of visual information processing is known as visual perception, an abnormality of which is called visual impairment, and a complete absence of which is called blindness. Non-image forming visual functions, independent of visual perception, include the pupillary light reflex and circadian photoentrainment.

In neuroanatomy, the lateral geniculate nucleus is a structure in the thalamus and a key component of the mammalian visual pathway. It is a small, ovoid, ventral projection of the thalamus where the thalamus connects with the optic nerve. There are two LGNs, one on the left and another on the right side of the thalamus. In humans, both LGNs have six layers of neurons alternating with optic fibers.
The receptive field, or sensory space, is a delimited medium where some physiological stimuli can evoke a sensory neuronal response in specific organisms.
Visual processing is a term that is used to refer to the brain's ability to use and interpret visual information from the world around us. The process of converting light energy into a meaningful image is a complex process that is facilitated by numerous brain structures and higher level cognitive processes. On an anatomical level, light energy first enters the eye through the cornea, where the light is bent. After passing through the cornea, light passes through the pupil and then lens of the eye, where it is bent to a greater degree and focused upon the retina. The retina is where a group of light-sensing cells, called photoreceptors are located. There are two types of photoreceptors: rods and cones. Rods are sensitive to dim light and cones are better able to transduce bright light. Photoreceptors connect to bipolar cells, which induce action potentials in retinal ganglion cells. These retinal ganglion cells form a bundle at the optic disc, which is a part of the optic nerve. The two optic nerves from each eye meet at the optic chiasm, where nerve fibers from each nasal retina cross which results in the right half of each eye's visual field being represented in the left hemisphere and the left half of each eye's visual fields being represented in the right hemisphere. The optic tract then diverges into two visual pathways, the geniculostriate pathway and the tectopulvinar pathway, which send visual information to the visual cortex of the occipital lobe for higher level processing.
The opponent process is a color theory that states that the human visual system interprets information about color by processing signals from photoreceptor cells in an antagonistic manner. The opponent-process theory suggests that there are three opponent channels, each comprising an opposing color pair: red versus green, blue versus yellow, and black versus white (luminance). The theory was first proposed in 1892 by the German physiologist Ewald Hering.
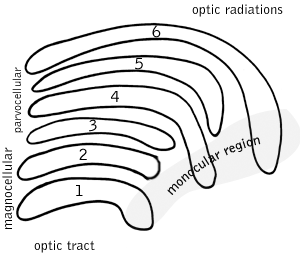
A koniocellular cell is a neuron with a small cell body that is located in the koniocellular layer of the lateral geniculate nucleus (LGN) in primates, including humans.

Retinotopy is the mapping of visual input from the retina to neurons, particularly those neurons within the visual stream. For clarity, 'retinotopy' can be replaced with 'retinal mapping', and 'retinotopic' with 'retinally mapped'.

In neurobiology, lateral inhibition is the capacity of an excited neuron to reduce the activity of its neighbors. Lateral inhibition disables the spreading of action potentials from excited neurons to neighboring neurons in the lateral direction. This creates a contrast in stimulation that allows increased sensory perception. It is also referred to as lateral antagonism and occurs primarily in visual processes, but also in tactile, auditory, and even olfactory processing. Cells that utilize lateral inhibition appear primarily in the cerebral cortex and thalamus and make up lateral inhibitory networks (LINs). Artificial lateral inhibition has been incorporated into artificial sensory systems, such as vision chips, hearing systems, and optical mice. An often under-appreciated point is that although lateral inhibition is visualised in a spatial sense, it is also thought to exist in what is known as "lateral inhibition across abstract dimensions." This refers to lateral inhibition between neurons that are not adjacent in a spatial sense, but in terms of modality of stimulus. This phenomenon is thought to aid in colour discrimination.

In vision, filling-in phenomena are those responsible for the completion of missing information across the physiological blind spot, and across natural and artificial scotomata. There is also evidence for similar mechanisms of completion in normal visual analysis. Classical demonstrations of perceptual filling-in involve filling in at the blind spot in monocular vision, and images stabilized on the retina either by means of special lenses, or under certain conditions of steady fixation. For example, naturally in monocular vision at the physiological blind spot, the percept is not a hole in the visual field, but the content is “filled-in” based on information from the surrounding visual field. When a textured stimulus is presented centered on but extending beyond the region of the blind spot, a continuous texture is perceived. This partially inferred percept is paradoxically considered more reliable than a percept based on external input..
A topographic map is the ordered projection of a sensory surface, like the retina or the skin, or an effector system, like the musculature, to one or more structures of the central nervous system. Topographic maps can be found in all sensory systems and in many motor systems.
Complex cells can be found in the primary visual cortex (V1), the secondary visual cortex (V2), and Brodmann area 19 (V3).

The colour centre is a region in the brain primarily responsible for visual perception and cortical processing of colour signals received by the eye, which ultimately results in colour vision. The colour centre in humans is thought to be located in the ventral occipital lobe as part of the visual system, in addition to other areas responsible for recognizing and processing specific visual stimuli, such as faces, words, and objects. Many functional magnetic resonance imaging (fMRI) studies in both humans and macaque monkeys have shown colour stimuli to activate multiple areas in the brain, including the fusiform gyrus and the lingual gyrus. These areas, as well as others identified as having a role in colour vision processing, are collectively labelled visual area 4 (V4). The exact mechanisms, location, and function of V4 are still being investigated.
A hypercomplex cell is a type of visual processing neuron in the mammalian cerebral cortex. Initially discovered by David Hubel and Torsten Wiesel in 1965, hypercomplex cells are defined by the property of end-stopping, which is a decrease in firing strength with increasingly larger stimuli. The sensitivity to stimulus length is accompanied by selectivity for the specific orientation, motion, and direction of stimuli. For example, a hypercomplex cell may only respond to a line at 45˚ that travels upward. Elongating the line would result in a proportionately weaker response. Ultimately, hypercomplex cells can provide a means for the brain to visually perceive corners and curves in the environment by identifying the ends of a given stimulus.
Globs are millimeter-sized color modules found beyond the visual area V2 in the brain's color processing ventral pathway. They are scattered throughout the posterior inferior temporal cortex in an area called the V4 complex. They are clustered by color preference, and organized as color columns. They are the first part of the brain in which color is processed in terms of the full range of hues found in color space.
Feature detection is a process by which the nervous system sorts or filters complex natural stimuli in order to extract behaviorally relevant cues that have a high probability of being associated with important objects or organisms in their environment, as opposed to irrelevant background or noise.

A parasol cell, sometimes called an M cell or M ganglion cell, is one type of retinal ganglion cell (RGC) located in the ganglion cell layer of the retina. These cells project to magnocellular cells in the lateral geniculate nucleus (LGN) as part of the magnocellular pathway in the visual system. They have large cell bodies as well as extensive branching dendrite networks and as such have large receptive fields. Relative to other RGCs, they have fast conduction velocities. While they do show clear center-surround antagonism, they receive no information about color. Parasol ganglion cells contribute information about the motion and depth of objects to the visual system.
Surround suppression is where the relative firing rate of a neuron may under certain conditions decrease when a particular stimulus is enlarged. It has been observed in electrophysiology studies of the brain and has been noted in many sensory neurons, most notably in the early visual system. Surround suppression is defined as a reduction in the activity of a neuron in response to a stimulus outside its classical receptive field.

Russell L. De Valois was an American scientist recognized for his pioneering research on spatial and color vision.

Laura Busse is a German neuroscientist and professor of Systemic Neuroscience within the Division of Neurobiology at the Ludwig Maximilian University of Munich. Busse's lab studies context-dependent visual processing in mouse models by performing large scale in vivo electrophysiological recordings in the thalamic and cortical circuits of awake and behaving mice.
References
- ↑ "Robert Shapley". Faculty Directory. Cns.nyu.edu. 2019-10-08. Retrieved 2022-10-07.
- ↑ "MacArthur Foundation".
- ↑ Hochstein, S. and Shapley, R. (1976) Linear and nonlinear spatial subunits in Y cat retinal ganglion cells. J.Physiol., 262, 265-284.
- ↑ Shapley, R. and Victor, J.D. (1978) The effect of contrast on the transfer properties of cat retinal ganglion cells, J.Physiol., 285, 275-298.
- ↑ Shapley R Kaplan E Soodak R. (1981) Spatial summation and contrast sensitivity of X and Y cells in the lateral geniculate nucleus of the macaque Nature 292, 543-545.
- ↑ Reid, R.C. and Shapley, R. (1992) Spatial structure of cone inputs to receptive fields in primate lateral geniculate nucleus, Nature, 356, 716-718.
- ↑ Reid RC and Shapley RM (2002) Space and time maps of cone photoreceptor signals in macaque lateral geniculate nucleus. J. Neurosci. 22:6158-6175.
- ↑ Ringach, D., Hawken, M. and Shapley, R. (1997) The dynamics of orientation tuning in the macaque monkey striate cortex, Nature, 387, 281-284.
- ↑ Johnson EA Hawken MJ and Shapley RM (2001) The Spatial Transformation of Color in the Primary Visual Cortex of the Macaque Monkey, Nature Neuroscience 4: 409-16.
- ↑ Johnson EN, Hawken MJ, Shapley R. (2004) Cone Inputs in Macaque Primary Visual Cortex. J Neurophysiol. 91:2501-14.
- ↑ Johnson EN, Hawken MJ, Shapley R. (2008) The orientation selectivity of color-responsive neurons in macaque V1. J Neurosci. 28:8096-8106.
- ↑ Burns SP, Xing D, Shelley MJ, Shapley RM (2010) Searching for autocoherence in the cortical network with a time-frequency analysis of the local field potential. J Neurosci. 30:4033-47.
- ↑ Burns SP, Xing D, Shapley RM. (2011) Is gamma-band activity in the local field potential of V1 cortex a "clock" or filtered noise? J Neurosci. 31:9658-64.
- ↑ Yeh CI, Xing D, Williams PE, Shapley RM. (2009) Stimulus ensemble and cortical layer determine V1 spatial receptive fields. Proc Natl Acad Sci U S A. 106:14652-7.
- ↑ Shapley, R. (1996) Guest editorial: Art and the perception of nature: Illusory contours in the paintings of Ellsworth Kelly, Perception, 25, 1259-1261.