A dolphin is an aquatic mammal in the clade Odontoceti. Dolphins belong to the families Delphinidae, Platanistidae, Iniidae, Pontoporiidae, and possibly extinct Lipotidae. There are 40 extant species named as dolphins.
Rapid eye movement sleep is a unique phase of sleep in mammals and birds, characterized by random rapid movement of the eyes, accompanied by low muscle tone throughout the body, and the propensity of the sleeper to dream vividly.

Cetacean intelligence is the overall intelligence and derived cognitive ability of aquatic mammals belonging in the infraorder Cetacea (cetaceans), including baleen whales, porpoises, and dolphins. In 2014, a study found for first time that the long-finned pilot whale has more neocortical neurons than any other mammal, including humans, examined to date.

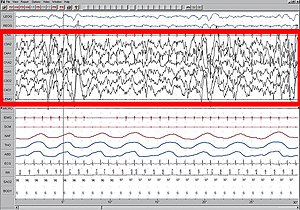
Delta waves are high amplitude neural oscillations with a frequency between 0.5 and 4 hertz. Delta waves, like other brain waves, can be recorded with electroencephalography (EEG) and are usually associated with the deep stage 3 of NREM sleep, also known as slow-wave sleep (SWS), and aid in characterizing the depth of sleep. Suppression of delta waves leads to inability of body rejuvenation, brain revitalization and poor sleep.

Cetacean surfacing behaviour is a grouping of movement types that cetaceans make at the water's surface in addition to breathing. Cetaceans have developed and use surface behaviours for many functions such as display, feeding and communication. All regularly observed members of the order Cetacea, including whales, dolphins and porpoises, show a range of surfacing behaviours.
Non-rapid eye movement sleep (NREM), also known as quiescent sleep, is, collectively, sleep stages 1–3, previously known as stages 1–4. Rapid eye movement sleep (REM) is not included. There are distinct electroencephalographic and other characteristics seen in each stage. Unlike REM sleep, there is usually little or no eye movement during these stages. Dreaming occurs during both sleep states, and muscles are not paralyzed as in REM sleep. People who do not go through the sleeping stages properly get stuck in NREM sleep, and because muscles are not paralyzed a person may be able to sleepwalk. According to studies, the mental activity that takes place during NREM sleep is believed to be thought-like, whereas REM sleep includes hallucinatory and bizarre content. NREM sleep is characteristic of dreamer-initiated friendliness, compared to REM sleep where it is more aggressive, implying that NREM is in charge of simulating friendly interactions. The mental activity that occurs in NREM and REM sleep is a result of two different mind generators, which also explains the difference in mental activity. In addition, there is a parasympathetic dominance during NREM. The reported differences between the REM and NREM activity are believed to arise from differences in the memory stages that occur during the two types of sleep.
Hypnagogia is the transitional state from wakefulness to sleep, also defined as the waning state of consciousness during the onset of sleep. Its opposite state is described as hypnopompia – the transitional state from sleep into wakefulness. Mental phenomena that may occur during this "threshold consciousness" phase include hypnagogic hallucinations, lucid dreaming, and sleep paralysis.
Sexsomnia, also known as sleep sex, is a distinct form of parasomnia, or an abnormal activity that occurs while an individual is asleep. Sexsomnia is characterized by an individual engaging in sexual acts while in non-rapid eye movement (NREM) sleep. Sexual behaviors that result from sexsomnia are not to be mistaken with normal nocturnal sexual behaviors, which do not occur during NREM sleep. Sexual behaviors that are viewed as normal during sleep and are accompanied by extensive research and documentation include nocturnal emissions, nocturnal erections, and sleep orgasms.

A K-complex is a waveform that may be seen on an electroencephalogram (EEG). It occurs during stage 2 NREM sleep. It is the "largest event in healthy human EEG". They are more frequent in the first sleep cycles.

The ventrolateral preoptic nucleus (VLPO), also known as the intermediate nucleus of the preoptic area (IPA), is a small cluster of neurons situated in the anterior hypothalamus, sitting just above and to the side of the optic chiasm in the brain of humans and other animals. The brain's sleep-promoting nuclei, together with the ascending arousal system which includes components in the brainstem, hypothalamus and basal forebrain, are the interconnected neural systems which control states of arousal, sleep, and transitions between these two states. The VLPO is active during sleep, particularly during non-rapid eye movement sleep, and releases inhibitory neurotransmitters, mainly GABA and galanin, which inhibit neurons of the ascending arousal system that are involved in wakefulness and arousal. The VLPO is in turn innervated by neurons from several components of the ascending arousal system. The VLPO is activated by the endogenous sleep-promoting substances adenosine and prostaglandin D2. The VLPO is inhibited during wakefulness by the arousal-inducing neurotransmitters norepinephrine and acetylcholine. The role of the VLPO in sleep and wakefulness, and its association with sleep disorders – particularly insomnia and narcolepsy – is a growing area of neuroscience research.

Slow-wave sleep (SWS), often referred to as deep sleep, is the third stage of non-rapid eye movement sleep (NREM), where electroencephalography activity is characterised by slow delta waves.

Part of the human brain, the basal forebrain structures are located in the forebrain to the front of and below the striatum. They include the ventral basal ganglia, nucleus basalis, diagonal band of Broca, substantia innominata, and the medial septal nucleus. These structures are important in the production of acetylcholine, which is then distributed widely throughout the brain. The basal forebrain is considered to be the major cholinergic output of the central nervous system (CNS) centred on the output of the nucleus basalis. The presence of non-cholinergic neurons projecting to the cortex have been found to act with the cholinergic neurons to dynamically modulate activity in the cortex.

Sleep in animals refers to a behavioral and physiological state characterized by altered consciousness, reduced responsiveness to external stimuli, and homeostatic regulation observed in various animals. Sleep has been observed in mammals, birds, reptiles, amphibians, and some fish, and, in some form, in insects and even in simpler animals such as nematodes. The internal circadian clock promotes sleep at night for diurnal organisms and in the day for nocturnal organisms. Sleep patterns vary widely among species. It appears to be a requirement for all mammals and most other animals.
Ponto-geniculo-occipital waves or PGO waves are distinctive wave forms of propagating activity between three key brain regions: the pons, lateral geniculate nucleus, and occipital lobe; specifically, they are phasic field potentials. These waves can be recorded from any of these three structures during and immediately before REM sleep. The waves begin as electrical pulses from the pons, then move to the lateral geniculate nucleus residing in the thalamus, and end in the primary visual cortex of the occipital lobe. The appearances of these waves are most prominent in the period right before REM sleep, albeit they have been recorded during wakefulness as well. They are theorized to be intricately involved with eye movement of both wake and sleep cycles in many different animals.
Sleep onset is the transition from wakefulness into sleep. Sleep onset usually transits into non-rapid eye movement sleep but under certain circumstances it is possible to transit from wakefulness directly into rapid eye movement sleep.

The relationship between sleep and memory has been studied since at least the early 19th century. Memory, the cognitive process of storing and retrieving past experiences, learning and recognition, is a product of brain plasticity, the structural changes within synapses that create associations between stimuli. Stimuli are encoded within milliseconds; however, the long-term maintenance of memories can take additional minutes, days, or even years to fully consolidate and become a stable memory that is accessible. Therefore, the formation of a specific memory occurs rapidly, but the evolution of a memory is often an ongoing process.

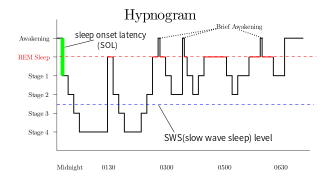
A hypnogram is a form of polysomnography; it is a graph that represents the stages of sleep as a function of time. It was developed as an easy way to present the recordings of the brain wave activity from an electroencephalogram (EEG) during a period of sleep. It allows the different stages of sleep: rapid eye movement sleep (REM) and non-rapid eye movement sleep (NREM) to be identified during the sleep cycle. NREM sleep can be further classified into NREM stage 1, 2 and 3. The previously considered 4th stage of NREM sleep has been included within stage 3; this stage is also called slow wave sleep (SWS) and is the deepest stage of sleep.

The neuroscience of sleep is the study of the neuroscientific and physiological basis of the nature of sleep and its functions. Traditionally, sleep has been studied as part of psychology and medicine. The study of sleep from a neuroscience perspective grew to prominence with advances in technology and the proliferation of neuroscience research from the second half of the twentieth century.
The parafacial zone (PZ) is a brain structure located in the brainstem within the medulla oblongata believed to be heavily responsible for non-rapid eye movement (non-REM) sleep regulation, specifically for inducing slow-wave sleep.
In birds, sleep consists of "periods of eye closure interrupted by short periods of eye-opening." During the short periods of eye-opening, electroencephalographic (EEG) studies indicate that the birds are still sleeping; the voltage level in the brain is identical. Birds restore their arousal thresholds during sleep. During their short eye-open periods, sleeping birds can mobilize almost instantaneously when threatened by a predator. Avian species have been found to rely on flock size and height for predatory precautions. Between the eye-opening and group sleep, these precautions allow sleep to be beneficial and safe.