Ascomycota is a phylum of the kingdom Fungi that, together with the Basidiomycota, forms the subkingdom Dikarya. Its members are commonly known as the sac fungi or ascomycetes. It is the largest phylum of Fungi, with over 64,000 species. The defining feature of this fungal group is the "ascus", a microscopic sexual structure in which nonmotile spores, called ascospores, are formed. However, some species of Ascomycota are asexual and thus do not form asci or ascospores. Familiar examples of sac fungi include morels, truffles, brewers' and bakers' yeast, dead man's fingers, and cup fungi. The fungal symbionts in the majority of lichens such as Cladonia belong to the Ascomycota.

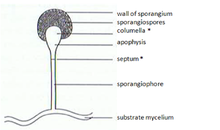
A sporangium ; pl.: sporangia) is an enclosure in which spores are formed. It can be composed of a single cell or can be multicellular. Virtually all plants, fungi, and many other groups form sporangia at some point in their life cycle. Sporangia can produce spores by mitosis, but in land plants and many fungi, sporangia produce genetically distinct haploid spores by meiosis.
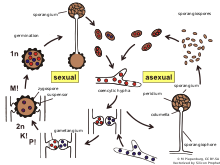
A zygospore is a diploid reproductive stage in the life cycle of many fungi and protists. Zygospores are created by the nuclear fusion of haploid cells. In fungi, zygospores are formed in zygosporangia after the fusion of specialized budding structures, from mycelia of the same or different mating types, and may be chlamydospores. In many eukaryotic algae, including many species of the Chlorophyta, zygospores are formed by the fusion of unicellular gametes of different mating types.

Phycomyces blakesleeanus is a filamentous fungus in the Order Mucorales of the phylum Zygomycota or subphylum Mucoromycotina. The spore-bearing sporangiophores of Phycomyces are very sensitive to different environmental signals including light, gravity, wind, chemicals and adjacent objects. They exhibit phototropic growth: most Phycomyces research has focused on sporangiophore photobiology, such as phototropism and photomecism. Metabolic, developmental, and photoresponse mutants have been isolated, some of which have been genetically mapped. At least ten different genes are required for phototropism. The madA gene encodes a protein related to the White Collar-1 class of photoreceptors that are present in other fungi, while madB encodes a protein related to the White Collar-2 protein that physically bind to White collar 1 to participate in the responses to light.

Phycomyces is a genus of fungus in the Zygomycota phylum. They are known for their strong phototropism response and helical growth of the sporangium. The best studied species is Phycomyces blakesleeanus.

The Mucorales is the largest and best-studied order of zygomycete fungi. Members of this order are sometimes called pin molds. The term mucormycosis is now preferred for infections caused by molds belonging to the order Mucorales.

Mucor is a microbial genus of approximately 40 species of molds in the family Mucoraceae. Species are commonly found in soil, digestive systems, plant surfaces, some cheeses like Tomme de Savoie, rotten vegetable matter and iron oxide residue in the biosorption process.
Sporogenesis is the production of spores in biology. The term is also used to refer to the process of reproduction via spores. Reproductive spores were found to be formed in eukaryotic organisms, such as plants, algae and fungi, during their normal reproductive life cycle. Dormant spores are formed, for example by certain fungi and algae, primarily in response to unfavorable growing conditions. Most eukaryotic spores are haploid and form through cell division, though some types are diploid sor dikaryons and form through cell fusion.we can also say this type of reproduction as single pollination

Blakeslea trispora is a mould and member of the division Zygomycota. This species has been well studied for its ability to produce carotenoids, particularly, β-carotene and lycopene. β-carotene is a vitamin A precursor and both of β-carotene and lycopene play a significant role in the inhibition of oxidative stress. Blakeslea trispora is commonly isolated from soil samples throughout the Southern United States and Southern Asia. B. trispora is a pathogen of tropical plants. In vivo pathogenicity testing using animal models suggests this fungus is not a cause of animal or human disease.
Mucor mucedo, commonly known as the common pinmould, is a fungal plant pathogen and member of the phylum Mucoromycota and the genus Mucor. Commonly found on soil, dung, water, plants and moist foods, Mucor mucedo is a saprotrophic fungus found world-wide with 85 known strains. It is often mistaken for Rhizopus rots on fruits due to similar mould growth shape and colour. Contrastingly, however, Mucor mucedo is found to grow on a wide range of stored grains and plants, including cucumber and tomato. Discovered in Italy in 1729 by P.A. Micheli and later noted by Carl Linnaeus in 1753 in the Species Plantarum, Mucor mucedo was originally classified as Mucor vulgaris by Micheli but later classified synonymous under name Mucor mucedo. The species was redescribed as Ascophora mucedo by H.J. Tode in 1790 but this type resided in a stoloniferous habitat and was later made the type of new genus Rhizopus.

Mucor racemosus is a rapidly growing, weedy mould belonging to the division Mucoromycota. It is one of the earliest fungi to be grown in pure culture and was first isolated in 1886. It has a worldwide distribution and colonizes many habitats such as vegetational products, soil and houses. The fungus is mostly known for its ability to exhibit both filamentous and yeast-like morphologies, often referred to as dimorphism. Stark differences are seen in both forms and conditions of the environment heavily affect the phases of the M. racemosus. Like many fungi, it also reproduces both sexually and asexually. The dimorphic capacity of this species has been proposed as an important factor in its pathogenicity and has enhanced the industrial importance. This species is considered an opportunistic pathogen, generally limited to immunocompromised individuals. It also been associated with allergy and inflammations of facial sinuses. Its association with allergy has made it a common fungus used in allergen medical testing. Industrial use of the fungus is in the production of enzymes and the manufacture of certain dairy foods.

The Cunninghamellaceae are a family of fungi in the order Mucorales.

The Zoopagomycotina are a subdivision of the fungal division Zygomycota sensu lato. It contains 5 families and 20 genera. Relationships among and within subphyla of Zygomycota are poorly understood, and their monophyly remains in question, so they are sometimes referred to by the informal name zygomycetes.

Spinellus fusiger, commonly known as bonnet mold, is a species of fungus in the phylum Mucoromycota. It is a pin mold that is characterized by erect sporangiophores that are simple in structure, brown or yellowish-brown in color, and with branched aerial filaments that bear the zygospores. It grows as a parasitic mold on mushrooms, including several species from the genera Mycena, including M. haematopus, M. pura, M. epipterygia, M. leptocephala, and various Collybia species, such as C. alkalivirens, C. luteifolia, C. dryophila, and C. butyracea. It has also been found growing on agaric species in Amanita, Gymnopus, and Hygrophorus.

Mortierella polycephala is a saprotrophic fungus with a wide geographical distribution occurring in many different habitats from soil and plants to salt marshes and slate slopes. It is the type species of the genus Mortierella, and was first described in 1863 by Henri Coemans. A characteristic feature of the fungus is the presence of stylospores, which are aerial, spiny resting spores (chlamydospores).

Syzygites is a monotypic genus in Zygomycota. The sole described species is Syzygites megalocarpus, which was the first fungus for which sex was reported and the main homothallic representative in the research that allowed for the classification of fungi as homothallic or heterothallic. It is also the fungus from which the term "zygospore" was coined.

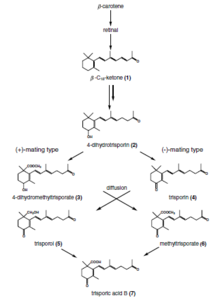
Trisporic acids (TSAs) are C-18 terpenoid compounds synthesized via β-carotene and retinol pathways in the zygomycetes. They are pheromone compound responsible for sexual differentiation in those fungal species. TSAs and related compounds make up the trisporoid group of chemicals.
Mucoromycota is a division within the kingdom fungi. It includes a diverse group of various molds, including the common bread molds Mucor and Rhizopus. It is a sister phylum to Dikarya.
Umbelopsis ramanniana is a common and abundant soil fungus. Although the ecological role of this fungus in natural ecosystems is not yet known, it is a cosmopolitan saprotroph in soil, forest leaf litter, in animal dung, and on the spore-producing bodies of ascomycete fungi. Umbelopsis ramanniana has also been found growing as an endophyte within xylem tissue of both healthy and declining conifers, but its exact effect on the plant hosts is unknown. Umbelopsis ramanniana is a representative of a unique group of zygomycete fungi that is distinct from the Mucoromycotina and Mortierellomycotina and instead forms an early diverging lineage within the Mucoralean fungi. Umbelopsis ramanniana is important from a biochemistry and biotechnology perspective because it is highly tolerant to fungicides of benomyl group, and it is oleaginous. Expression of Umbelopsis ramanniana diacylglycerol O-acyltransferase 2A in seed increases oil in soybean without reduction of other important yield parameters. This increase in oil can potentially add over $1 billion to the annual value of soybean crops.

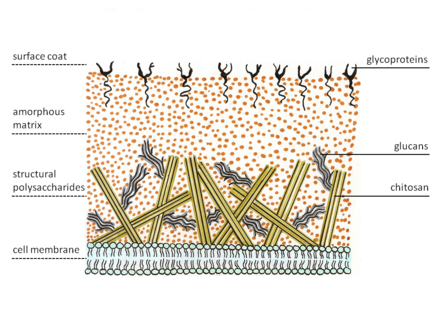
This glossary of mycology is a list of definitions of terms and concepts relevant to mycology, the study of fungi. Terms in common with other fields, if repeated here, generally focus on their mycology-specific meaning. Related terms can be found in glossary of biology and glossary of botany, among others. List of Latin and Greek words commonly used in systematic names and Botanical Latin may also be relevant, although some prefixes and suffixes very common in mycology are repeated here for clarity.