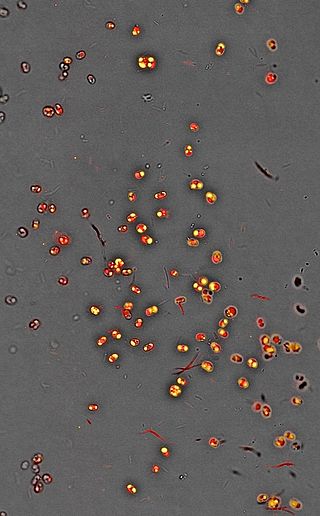
Chlamydomonas nivalis, also referred to as Chloromonas typhlos,[5][4] is a unicellular red-coloured photosyntheticgreen alga that is found in the snowfields of the alps and polar regions all over the world. They are one of the main algae responsible for causing the phenomenon of watermelon snow (also blood snow, raspberry snow[7]), where patches of snow appear red or pink. The first account of microbial communities that form red snow was made by Aristotle. Researchers have been active in studying this organism for over 100 years.
Although C. nivalis is closely related to Chlamydomonas reinhardtii, the environmental conditions each species inhabits are very different. C. nivalis can be found in mountains, snowfields, and polar regions around the world. The habitat of C. nivalis subjects the cells to environmental extremes including limited nutrients, low temperatures, and intense sunlight. In comparison with the mesophilicC. reinhardtii, C. nivalis has special mechanisms that allow it to be cryotolerant and survive on rock surfaces as well as in soil, meltwater, and snow. Secondary carotenoids, a thick cell wall, and particles on the cell wall are some characteristics that protect the cyst from light, drought, and radiation stress. Although the seasonal mobile to dormant life cycle of C. nivalis is complex, it also helps the algae exploit its niche and survive unfavourable conditions. As a result, C. nivalis is one of the best known and studied snow algae. When taking account of the photoprotective effect of its secondary carotenoid, astaxanthin, among the other adaptive mechanisms to its extreme habitat, it can be understood how C. nivalis became so dominant in microbial snow algae communities. Green motile offspring are produced in the spring and throughout the summer. They develop into red dormant cysts, the stage where this organism spends most of its life cycle, as the winter season begins and remain a cyst until the spring.
This alga is an interesting organism for researchers in various fields to study due to its possible role in lowering global albedo, ability to survive in extreme environments, and production of commercially relevant compounds. Additionally, its life cycle is still being studied today in an effort to better understand this organism and amend previous classification errors.
Etymology
The name Chlamydomonas nivalis is of compound Greek and Latin origin. Chlamydomonas is ultimately derived from the Ancient Greek χλαμύς (khlamús, “cloak, mantle”) and μονάς (monás, “solitary”),[8] while nivalis, from the Latinnivālis, translates to ‘found growing in or near snow’, as this species of algae are only found associated with snow or near snowy areas.[9][10]
Description
The seasonal life cycle of C. nivalis can be broken down to three stages based on the colour of the cell as a result of carotenoid composition, which are green, orange, and red.[11] Orange cells and red cells are the most difficult to differentiate as they look similar while the red and green cells are easiest to differentiate as they have more significant differences in composition.[12] Cells at the red stage were previously described as a separate species than the green cells, but were later discovered to be different stages of the C. nivalis’ complex life cycle.[2]
Small green coloured motile cells of the young C. nivalis at the green stage are produced in spring or early summer when temperatures are warmer and zygotes undergo meiosis in meltwater pools.[13] The biflagellated cells are slightly oval and about 5-15µm in diameter.[12][14] In this asexually reproductive phase, the cells are sensitive to temperature and drought stress. They avoid unfavourable light and temperature by swimming in the snow until they reach more optimal conditions.[15][13][11]Chloroplasts of green cells are irregularly shaped.[11] The dominating pigment, chlorophyll, gives the cell its characteristic hue and facilitates maximum cell growth through light absorption. Secondary carotenoid concentrations are much lower at this stage as the cells need photosynthetically active radiation for energy and growth.[12] Cells in the green stage also have less organic and inorganic particles on their surface compared to mature cysts.[15]
Later in the season, when nitrogen and nutrients becomes limited and radiation stress increases, the green cells will develop into flagellated sexual gametes that mate and produce new zygotes that have lost their flagella and are capable of surviving the winter period.[16][14] Transformation into the zygote, or hypnoblast, is characterized by the production and accumulation of reserve materials that include sugars and lipids as well as the formation of esterified secondary carotenoids.[15] The secondary carotenoids will turn the green zygotes orange as they accumulate in the area around the plastids of the cell to protect the zygotes from UV radiation.[15][12] Orange and red spores can be seen throughout the summer. During this stage, the cell wall will also begin to thicken to help the cell tolerate freezing temperatures and UV light.[14][17] In addition, the color of these pigments reduces albedo such that individual cells may melt nearby ice and snow crystals to access limiting nutrients and water in an otherwise unavailable frozen state.[18]
History
The earliest documentation of red snow was made by Aristotle.[19] While he recognized that something must be contributing to the odd colouration, red snow was also commonly mistaken as mineral deposits or pollen up until the early 1900s.[20] In 1819, samples of ‘red snow’ were brought back for examination with a returning Arctic expedition under Sir John Ross. The samples were sent to Robert Brown and Francis Bauer for examination. Both men came to different conclusions on how to classify the specimens. Brown believed the specimen to be a unicellular alga while Bauer declared it a new species of fungus, Uredo nivalis.[20][1][21] Over the next century, many researchers disputed over whether these organisms were lichen, plants, alga, or animal. It was not until the early 20th century when researchers finally began to agree on the algal nature of the organism and gave its currently known name, Chlamydomonas nivalis.[1][21] In 1968 C. nivalis was officially recognized as a collective taxon.[22] Unfortunately, due to the lack of sequencing techniques, reliance on visually examining similarly looking snow alga, and complicated life cycle of this species, errors continued to be made in classifying this and other species of snow algae. Today, C. nivalis has become one of the most well-studied snow algae. Although its taxonomy is still being settled, the life cycle of this snow algae is now much better understood.[23][1][2][24] The historical disputes about the classification and misclassification of specimens have resulted in a number of names from older publications that all mean to refer to C. nivalis. These are: Uredo nivalis, Sphaerella nivalis, Protococcus nivalis, and Haematococcus nivalis.[1][2][3]
Habitat and ecology
C. nivalis has been reported worldwide in mountainous regions, polar regions, or snowfields of every continent.[23][25] It is the most abundant snow algae and typically composes the majority of cells identified in specimens taken from various sample sites.[15] Most habitats these algae reside in are very different from other species of the rest of the genus Chlamydomonas.[16] This includes, but is not limited to snow, rock surfaces, soil, meltwater, and cryoconite holes.[26][27][16][28]
The environmental conditions C. nivalis is typically exposed to are considered to be extreme. The cells can experience low nutrient availability, acidity, intense sunlight, radiation, extreme temperature regimes, and darkness.[23][13][26][29] Red-snow algae have been shown experimentally to be limited by both nutrients (N, P, and K) and liquid water.[30]C. nivalis spends the majority of its life in the cyst stage surrounded by snow at a depth that can range from 0–20 centimetres (0.0–7.9in).[14][31] This can change depending on if the cell is in a mobile stage and can move, the snow melts due to the onset of warm weather, or the onset of precipitation causes more snow to fall on the cells.[26] Cells that are exposed on unshaded snow may be subjected to high levels of visible light and ultraviolet radiation for an extended amount of time. Meanwhile, cells that are deep below the snow’s surface may experience darkness.[23] In its flagellated stage, the cell can move until it is in the most optimal position in the snow for moisture content, light, and temperature.[15] When in the immotile cyst stage, the C. nivalis cells must depend on the flow of meltwater to move it by chance to a favourable area.[26]
The temperatures in which this species can survive in ranges from below 0°C to just above 20°C. Growth is slow when temperatures are below 5°C. At 5-15°C the growth of C. nivalis cells can outperform the growth of C. reinhardtii cells.[27] Both species grow at the same rate at 20-25°C. The growth of C. nivalis is suppressed when temperatures rise above 30°C.[27] It is a true snow alga because it performs better in low temperatures than warm temperatures.[15] Due to C. nivalis’ ability to perform photosynthesis well from cold to moderate temperatures, this species is considered a cryotolerant mesophile rather than a cryophile.[3][27] This organism is also very resilient as they can also survive in warm soil for weeks. They can also tolerate dryness and room temperature for as long as 6 months.[15]
Fungi, worms, bacteria, and viruses have been found to associate with or live in the same environment as C. nivalis. Encapsulatedrod-shapedgram-negative bacteria have been found on the surface of C. nivalis cysts. The unknown bacteria were not detected in control samples that did not contain C. nivalis which strongly suggests that it must be associated with the algae.[32] Another bacterium, Mesorhizobium loti, was found as contamination in a C. nivalis culture, but further testing suggested that this bacteria may be synthesizing vitamin B12 for the algae.[33] In cryoconite holes C. nivalis can be found among bacteria, virus-like particles, ciliates, and Chlorophyte species.[28]Ice worms have also been found to live preferentially under C. nivalis in glaciers, possibly using the algae as a food source.[34] Infections of C. nivalis cells by chytrids, Chytridium chlamydococci, filamentous fungi, and Selenotila nivalis have also been observed.[3]
As winter approaches, the cells will approach the last stage of their life cycle. The orange cells mature into red cysts, the form in which it will remain for the remainder and longest portion of its life cycle.[26][35][14] Cells at this stage are most resistant to harsh environmental conditions.[15] Inorganic and organic materials such as bacteria, fungi, and dust particles coat the mucilage layer of the cell wall.[15] The inorganic impurities were found to be rich in silicon, iron, and aluminum. These elements can also be taken up into the cellular compartment and stored in vacuoles and may be an important source of mineral supply.[36] The cell wall, as the boundary that protects the inner contents of the cell from the harsh conditions in its habitat, is very rigid and hard to destroy.[15] It also may play a role in protecting the algal cells from desiccation during the freeze-thaw cycle alternations during seasonal changes.[32] The spherical immotile red cysts range from 35-40µm in diameter. The cell contains one central chloroplast that has a naked pyrenoid, ribosomes, starch grains, and numerous small grana stacks composed of 3-7 thylakoids within it.[23][15][32][37] Negatively charged phosphatidylglycerol composes the majority of the thylakoid membranes.[23] The thylakoid membrane lipid composition can also be changed to enhance lipid fluidity in response to lower temperatures.[27] An undulated membrane encloses the chloroplast. Lipid bodies and carotenoid globules surround the plastid.[37] A red secondary pigment, astaxanthin and esterified derivatives of it, accumulates up to 20 times the amount of chlorophyll a in the cytoplasmic lipid bodies of mature red spores.[23][15] Astaxanthin protects the chloroplast from excessive light by absorbing a portion of it before it reaches the photosynthetic apparatus which subsequently prevents photoinhibition and UV damage.[35] The absorbed radiation is converted to heat, aiding in the melt of nearby snow and ice crystals to access needed nutrients and liquid water.[18] Astaxanthin can also act as a metabolic sink for the metabolically active spores that do not divide.[23][17]
Within the cytoplasm there are several small cytoplasmic vacuoles with partially crystallized content within it.[36] While mitochondria are present, they are not very obvious. Most of the cytoplasmic space is taken up by the large plastid, lipid bodies, and carotenoid globules.[15][32]C. nivalis has one centrally located nucleus that is also oriented such that it is covered by the carotenoid globules full of astaxanthin that will provide protection against UV radiation.[26] The majority (91%) of astaxanthin derivatives are stored in its monoester form within dormant C. nivalis red cysts.[23][13] Astaxanthin is the pigment that makes the cell appear deep red. Other pigments that can also be found in C. nivalis include violaxanthin and adonirubin.[17]
Role in environmental processes and research
Visible algal blooms could be a crucial determinant of surface albedo.[38] It has been suggested that algal blooms partially composed of C. nivalis may contribute to lowering ice and snow albedo.[38] The red coloured pigments produced by the cell in combination with inorganic material could enhance the darkening over the snow and reduce the surface area of white snow.[39] Due to the absorption of solar energy by the alga, albedo would be reduced and the darker areas on the snow where the blooms form would melt more rapidly.[39] As a result, populations of C. nivalis would increase, creating a feedback loop that amplifies melting and reduces sunlight absorbance which contributes to glacier retreat and lowering albedo, as shown experimentally.[30] This is concerning to environmentalists and climate scientists.[40][41][42]
C. nivalis can be used as a model species for studying the cellular response mechanism to stressful conditions given the harsh conditions of its habitat.[43] It is also an important organism to study adaptation to extreme environments and may become one of the leading systems for research in cold adaptation.[2]C. nivalis is likely to have strong antioxidant capabilities, a robust repair mechanism, and other components that may be of interest to researchers.[26]
Thermophilicmicroalgae have gained biotechnological interest as a source for thermostable enzymes and commercial interest as a source for astaxanthin.[44]C. nivalis could also potentially be a source for pharmaceuticals, supplements, or beauty products if the algae could be mass produced for its astaxanthin.[45][46][47] The snow algae itself is likely safe to eat as there is no evidence supporting that it would cause diarrhea when ingested.[48]
Related Research Articles
Chlamydomonas is a genus of green algae consisting of about 150 species of unicellular flagellates, found in stagnant water and on damp soil, in freshwater, seawater, and even in snow as "snow algae". Chlamydomonas is used as a model organism for molecular biology, especially studies of flagellar motility and chloroplast dynamics, biogenesis, and genetics. One of the many striking features of Chlamydomonas is that it contains ion channels (channelrhodopsins) that are directly activated by light. Some regulatory systems of Chlamydomonas are more complex than their homologs in Gymnosperms, with evolutionarily related regulatory proteins being larger and containing additional domains.
Chlamydomonas reinhardtii is a single-cell green alga about 10 micrometres in diameter that swims with two flagella. It has a cell wall made of hydroxyproline-rich glycoproteins, a large cup-shaped chloroplast, a large pyrenoid, and an eyespot that senses light.
Astaxanthin is a keto-carotenoid within a group of chemical compounds known as terpenes. Astaxanthin is a metabolite of zeaxanthin and canthaxanthin, containing both hydroxyl and ketone functional groups. It is a lipid-soluble pigment with red coloring properties, which result from the extended chain of conjugated double bonds at the center of the compound.
Canthaxanthin is a keto-carotenoid pigment widely distributed in nature. Carotenoids belong to a larger class of phytochemicals known as terpenoids. The chemical formula of canthaxanthin is C40H52O2. It was first isolated in edible mushrooms. It has also been found in green algae, bacteria, crustaceans, and bioaccumulates in fish such as carp, golden grey mullet, seabream and trush wrasse.
Ice algae are any of the various types of algal communities found in annual and multi-year sea or terrestrial ice. On sea ice in the polar oceans, ice algae communities play an important role in primary production. The timing of blooms of the algae is especially important for supporting higher trophic levels at times of the year when light is low and ice cover still exists. Sea ice algal communities are mostly concentrated in the bottom layer of the ice, but can also occur in brine channels within the ice, in melt ponds, and on the surface.
Dunaliella salina is a type of halophile green unicellular micro-algae especially found in hypersaline environments, such as salt lakes and salt evaporation ponds. Known for its antioxidant activity because of its ability to create large amount of carotenoids, it is responsible for most of the primary production in hypersaline environments worldwide, and is also used in cosmetics and dietary supplements.
Watermelon snow, also called snow algae, pink snow, red snow, or blood snow, is a phenomenon caused by Chlamydomonas nivalis, a species of green algae containing a secondary red carotenoid pigment (astaxanthin) in addition to chlorophyll. Unlike most species of fresh-water algae, this species appears to be cryophilic (cold-loving) and thrives in freezing water.
Biological pigments, also known simply as pigments or biochromes, are substances produced by living organisms that have a color resulting from selective color absorption. Biological pigments include plant pigments and flower pigments. Many biological structures, such as skin, eyes, feathers, fur and hair contain pigments such as melanin in specialized cells called chromatophores. In some species, pigments accrue over very long periods during an individual's lifespan.
The eyespot apparatus is a photoreceptive organelle found in the flagellate or (motile) cells of green algae and other unicellular photosynthetic organisms such as euglenids. It allows the cells to sense light direction and intensity and respond to it, prompting the organism to either swim towards the light, or away from it. A related response occurs when cells are briefly exposed to high light intensity, causing the cell to stop, briefly swim backwards, then change swimming direction. Eyespot-mediated light perception helps the cells in finding an environment with optimal light conditions for photosynthesis. Eyespots are the simplest and most common "eyes" found in nature, composed of photoreceptors and areas of bright orange-red red pigment granules. Signals relayed from the eyespot photoreceptors result in alteration of the beating pattern of the flagella, generating a phototactic response.
Scenedesmus is a genus of green algae, in the class Chlorophyceae. They are colonial and non-motile.
Dunaliella is a single-celled, photosynthetic green alga, that is characteristic for its ability to outcompete other organisms and thrive in hypersaline environments. It is mostly a marine organism, though there are a few freshwater species that tend to be more rare. It is a genus in which certain species can accumulate relatively large amounts of β-carotenoids and glycerol in very harsh growth conditions consisting of high light intensities, high salt concentrations, and limited oxygen and nitrogen levels, yet is still very abundant in lakes and lagoons all around the world.
Chloromonas is a genus of green algae in the family Chlamydomonadaceae. It is closely related to the model green algae, Chlamydomonas, and distinguished mainly through the absence of a pyrenoid.
Trebouxia is a unicellular green alga. It is a photosynthetic organism that can exist in almost all habitats found in polar, tropical, and temperate regions. It can either exist in a symbiotic relationship with fungi in the form of lichen or it can survive independently as a free-living organism alone or in colonies. Trebouxia is the most common photobiont in extant lichens. It is a primary producer of marine, freshwater and terrestrial ecosystems. It uses carotenoids and chlorophyll a and b to harvest energy from the sun and provide nutrients to various animals and insects.
Haematococcus pluvialis is a freshwater species of Chlorophyta from the family Haematococcaceae. This species is well known for its high content of the strong antioxidant astaxanthin, which is important in aquaculture, and cosmetics. The high amount of astaxanthin is present in the resting cells, which are produced and rapidly accumulated when the environmental conditions become unfavorable for normal cell growth. Examples of such conditions include bright light, high salinity, and low availability of nutrients. Haematococcus pluvialis is usually found in temperate regions around the world. Their resting cysts are often responsible for the blood-red colour seen in the bottom of dried out rock pools and bird baths. This colour is caused by astaxanthin which is believed to protect the resting cysts from the detrimental effect of UV-radiation, when exposed to direct sunlight.
Phototaxis is a kind of taxis, or locomotory movement, that occurs when a whole organism moves towards or away from a stimulus of light. This is advantageous for phototrophic organisms as they can orient themselves most efficiently to receive light for photosynthesis. Phototaxis is called positive if the movement is in the direction of increasing light intensity and negative if the direction is opposite.
Nannochloropsis is a genus of algae comprising six known species. The genus in the current taxonomic classification was first termed by Hibberd (1981). The species have mostly been known from the marine environment but also occur in fresh and brackish water. All of the species are small, nonmotile spheres which do not express any distinct morphological features that can be distinguished by either light or electron microscopy. The characterisation is mostly done by rbcL gene and 18S rRNA sequence analysis.
Nannochloropsis is a genus of alga within the heterokont line of eukaryotes, that is being investigated for biofuel production. One marine Nannochloropsis species has been shown to be suitable for algal biofuel production due to its ease of growth and high oil content, mainly unsaturated fatty acids and a significant percentage of palmitic acid. It also contains enough unsaturated fatty acid linolenic acid and polyunsaturated acid for a quality biodiesel.
Euglena sanguinea is a species of the genus Euglena. The red colour is due to the presence of astaxanthin and the cells can be populous enough to colour water red. The pigment is used to protect the chloroplasts from light that is too intense, but as the light levels change the cells can take on a green colour as the red pigment is moved to the centre of the cells. Euglena sanguinea is known to make the potent icthyotoxin euglenophycin.
Sammy Boussiba is a professor emeritus at the French Associates Institute for Agriculture and Biotechnology of Drylands at the Jacob Blaustein Institutes for Desert Research at Ben-Gurion University of the Negev, Israel.
Protists are the eukaryotes that cannot be classified as plants, fungi or animals. They are mostly unicellular and microscopic. Many unicellular protists, particularly protozoans, are motile and can generate movement using flagella, cilia or pseudopods. Cells which use flagella for movement are usually referred to as flagellates, cells which use cilia are usually referred to as ciliates, and cells which use pseudopods are usually referred to as amoeba or amoeboids. Other protists are not motile, and consequently have no built-in movement mechanism.
References
1 2 3 4 5 Sutton, E. A. (1970). “The physiology and life histories of selected cryophytes of the pacific Northwest”. Ph.D. Thesis. Oregon state university, Corvallis.
1 2 3 4 5 Cvetkovska, M. C.; Hüner, N. P. A.; Smith, D. R. (2016). “Chilling out: the evolution and diversification of psychrophilic algae with a focus on Chlamydomonas”. Polar Biol. 40 (6): 1169-1184. doi: 10.1007/s00300-016-2045-4
1 2 3 4 Cepak, V.; Lukavsky, J. (2013) “Cryoseston of the Pirin Mountains, Bulgaria”. Acta Bot Croat. 72 (2): 257-268. doi: 10.2478/botcro-2013-0012.
↑ Guiry, M. D. in Guiry, M.D. & Guiry, G.M. (2018). AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; Accessed on: 04 March 2018.
1 2 3 Lütz, C. (2012). “Plants in Alpine Regions”. Springer-Verlag Wien. doi: 10.1007/978-3-7091-0136-0
1 2 3 4 Beer, T.; Tanaka, Z.; Netzter, N.; Rothschild, L. J.; Chen, B. (2011). “An analysis of uncultured extremophilic snow algae by non-invasive single cell Raman spectroscopy”. Proc. SPIE 8152, Instruments, Methods, and Missions for Astrobiology XIV, 81520F. doi: 10.1117/12.896481
1 2 3 4 Bidigare, R. R.; Ondrusek, M. E.; Kennicutt II, M. C.; Iturriaga, R.; Harvey, H. R.; Hoham, H. W.; Macko, S. A. (1993). “Evidence for a photoprotective function for secondary carotenoids of snow algae”. J. Phycol. 29 (4): 427-434.
1 2 3 4 5 Stibal, M.; Elster, J.; Sabacka, M.; Kastovska, K. (2007). “Seasonal and diel changes in photosynthetic activity of the snow alga Chlamydomonas nivalis (Chlorophyceae) from Svalbard determined by pulse amplitude modulation fluorometry”. FEMS Microbiology Ecology. 59 (2): 265–273. doi: 10.1111/j.1574-6941.2006.00264.x.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 Remias, D.; Lütz-Meindl, U.; Lütz C. (2005). “Photosynthesis pigments and ultrastructure of the alpine snow alga Clamydomonas nivalis”. European Journal of Phycology. 40 (3): 259-268, doi: 10.1080/09670260500202148
1 2 3 Müller, T.; Bleiss, W.; Martin, C.-D.; Rogaschewski, S.; Fuhr, G. (1998). “Snow algae from northwest Svalbard: their identification, distribution, pigment and nutrient content”. Polar Biology. 20 (1): 14-32.
1 2 Dial, R.; Ganey, G.; Skiles, M. (2018). “What color should glacier algae be? An ecological role for red carbon in the cryosphere”. FEMS Microbiology Ecology. 94 (3): fiy007. https://academic.oup.com/femsec/article/94/3/fiy007/4810544; Accessed on: 18 December 2018
1 2 Clark, F. C. (1875) “Red Snow”. Am. Nat. 9:129–135.
1 2 Wille, N. (1903). “Algologische Notizen IX-XIV. Nytt” Magazin for Naturvidenskaberne. 41: 89-185.
↑ Kol, E. (1968). “A note on red snow from New Zealand”. N. Z. J. Bot. 6 (2): 243-244. doi:10.1080/0028825X.1968.10429063
1 2 3 4 5 6 7 8 9 Remias, D.; Pichrtova, M.; Pangratz, M.; Lütz, C.; Holzinger, A. (2016) “Ecophysiology, secondary pigments and ultrastructure of Chlainomonas sp. (Chlorophyta) from the European Alps compared with Chlamydomonas nivalis forming red snow”. FEMS Microbiol. Ecol. 92 (4). doi: 10.1093/femsec/fiw030
↑ Brown, S. P.; Olson, B. J. S. C.; Jumpponen, A. (2015). “Fungi and algae co-occur in snow: an issue of shared habitat or algal facilitation of heterotrophs?”. Arct. Antarct. Alpine Res. 47 (4): 729–749. doi: 10.1657/AAAR0014-071
↑ Uetake, J.; Yoshimura, Y.; Nagatsuka, N.; Kanda, H. (2012). “Isolation of oligotrophic yeasts from supraglacial environments of different altitude on the Gulkana Glacier (Alaska)”. FEMS Microbiol Ecol 82 (2): 279–286. doi: 10.1111/j.1574-6941.2012.01323.x.
1 2 3 4 5 6 7 Gorton, H. L.; Vogelmann, T. C. (2003). “Ultraviolet radiation and the snow alga Chlamydomonas nivalis(Bauer) Wille”. Photochemistry and Photobiology. 77 (6): 608-615. doi: 10.1562/0031-8655(2003)0770608URATSA2.0.CO2
1 2 3 4 5 Lukes, M.; Prochazkova, L.; Shmidt, V.; Nedbalova, L.; Kaftan, D. (2014). “Temperature dependence of photosynthesis and thylakoid lipid composition in the red snow alga Chlamydomonas cf. nivalis (Chlorophyceae)”. FEMS Microbiol. Ecol. 89 (2): 303-315. doi: 10.1111/1574-6941.12299
1 2 Säwström, C.; Mumford, P.; Marshall, W.; Hodson, A.; Laybourn-Parry, J. (2002). “The microbial communities and primary productivity of cryoconite holes in an Arctic glacier (Svalbard 79°N)”. Polar Biology. 25 (8): 591-596.
↑ Leya, T.; Müller, T.; Ling, H. U.; Fuhr, G. (2004). “Snow algae from north-western Spitsbergen (Svalbard), Ber. Polarforsch. Meeresforsch. 492: 46-54.
1 2 Ganey, G.Q.; Loso, M.; Bryant Burgess, A.; Dial, R.J. (2017). “The role of microbes in snowmelt and radiative forcing on an Alaskan icefield”. Nature Geoscience. 10: 754-759. doi: 10.1038/NGEO3027
↑ Mosser, J. L.; Mosser, A. G.; Brock. T. D. (1977). “Photosynthesis in the snow: the alga Chlamydomonas nivalis (Chlorophyceae)”. J. Phycol. 13 (1): 22-27. doi: 10.1111/j.1529-8817.1977.tb02881.x
1 2 3 4 Weiss, R. L. (1983). “Fine structure of the snow alga (Chlamydomonas nivalis) and associated bacteria”. J. Phycol. 19 (2): 200-204. doi: 10.1111/j.0022-3646.1983.00200.x
↑ Kazamia, E.; Czesnick, H.; Nguyen, T. T.; Croft, M. T.; Sherwood, E.; Sasso, S.; Hodson, S. J.; Warren, M. J.; Smith, A. G. (2012). “Mutualistic interactions between vitamin B12 –dependent algae and heterotrophic bacteria exhibit regulation”. Environ. Microbiol. 14 (6): 1466-1476. doi: 10.1111/j.1462-2920.2012.02733.x.
↑ Shain, D. H.; Mason, T. A.; Farrell, A. H.; Michalewicz, L. A. (2001). “Distribution and behaviour of ice worms (Mesenchytraeus solifugus) in south-central Alaska”. Canadian Journal of Zoology. 79 (10): 1813-1821. doi: 10.1139/z01-143
1 2 Williams, W. E.; Gorton, H. L.; Vogelmann, T. C. (2003). “Surface gas-exchange processes of snow algae”. Proc. Natl. Acad. Sci. USA. 100 (2): 562-566. doi: 10.1073/pnas.0235560100
1 2 Lütz-Meindl, U.; Lütz, C. (2006). “Analysis of element accumulation in cell wall attached and intracellular particles of snow algae by EELS and ESI”. Micron. 37 (5): 452-458.
1 2 Holzinger, A.; Lutz, C. (2006). “Algae and UV irradiation: Effects on ultrastructure and related metabolic functions”. Micron 37 (3): 190–207. doi: 10.1016/j.micron.2005.10.015.
1 2 Cook. J. M.; Hodson, A. J.; Taggart, A. J.; Mernild, S. H.; Tranter, M. (2017). “A predictive model for the spectral “bioalbedo” of 30 snow”. J. Geophys. Res. Earth Surf. 122 (1). doi:10.1002/2016JF003932, 2017.
1 2 Cook, J.M.; Hodson, A.; Gardner, A. S.; Flanner, M.; Tedstone, A. J.; Williamson, C.; et al (2017). “Quantifying bioalbedo: a new physically based model and discussion of empirical methods for characterising biological influence on ice and snow albedo”. The Cryosphere. 11: 2611-2632. doi: 10.5194/tc-11-2611-2017
↑ Thomas, W. H.; Duval, B. (1995). “Sierra Nevada, California, U.S.A., snow algae: snow albedo changes, algal-bacterial interrelationships, and ultraviolet radiation effects”. Arct. Alp. Res. 27 (4): 389-399. doi: 10.2307/1552032
↑ Hisakawa, N.; Quistad, S.D.; Hester, E.R.; Martynova, D.; Maughan, H.; Sala, E.; Rohwer, F. (2015). “Metagenomic and satellite analyses of red snow in the Russian Arctic”. PeerJ. 3, doi: 10.7717/peerj.1491.
↑ Lutz, S.; Anesio, A. M.; Raiswell, R.; Edwards, A.; Newton, R. J.; Gill, F.; et al. (2016). “The biogeography of red snow microbiomes and their role in melting arctic glaciers”. Nat. Commun. 7. doi:11968 10.1038/ncomms11968
↑ Wiencke, C.; Clayton, M.N. (2009). “Biology of polar benthic algae”. Bot. Mar. 52: 479–481. doi: 10.1515/BOT.2009.083
↑ Varshney, P.; Mikulic, P.; Vonshak, A.; Beardall, J.; Wangikar, P.P. (2015). “Extremophilic micro-algae and their potential contribution in biotechnology”. Bioresour. Technol. 184: 363–372. doi: 10.1016/j.biortech.2014.11.040
↑ Duval, B.; Shetty, K.; Thomas, W. H. (1999). “Phenolic compounds and antioxidant properties in the snow alga Chlamydomonas nivalis after exposure to UV light”. J. Appl. Phycol. 11: 559-566. doi: 10.1023/A:1008178208949
↑ Martin, J. F.; Gudina, E.; Barredo, J. L. (2008). “Conversion of beta-carotene into astaxanthin: Two separate enzymes or a bifunctional hydroxylase-ketolase protein?”. Mirob. Cell. Fact. 20 (7): 3. doi: 10.1186/1475-2859-7-3.
↑ Tominaga, M.; Beinlich, A.; Lima, E. A.; Tivey, M. A.; Hampton, B. A.; Weiss, B.; Harigane, Y. (2017). “Multi-scale magnetic mapping of serpentinite carbonation”. Nature Communications. 8: 1870-1880. doi:10.1038/s41467-017-01610-4
↑ Fiore, D. C.; Mckee, D. D.; Janiga, M. A. (1997). “Red snow: is it safe to eat? A pilot study”. Wilderness Environ. Med. 8 (2): 94-95. doi: 10.1580/1080-6032(1997)008[0094:RSIIST] 2.3.CO;2
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.