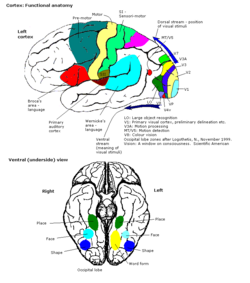
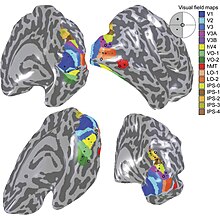
The visual cortex of the brain is the area of the cerebral cortex that processes visual information. It is located in the occipital lobe. Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex. The area of the visual cortex that receives the sensory input from the lateral geniculate nucleus is the primary visual cortex, also known as visual area 1 (V1), Brodmann area 17, or the striate cortex. The extrastriate areas consist of visual areas 2, 3, 4, and 5.

Color constancy is an example of subjective constancy and a feature of the human color perception system which ensures that the perceived color of objects remains relatively constant under varying illumination conditions. A green apple for instance looks green to us at midday, when the main illumination is white sunlight, and also at sunset, when the main illumination is red. This helps us identify objects.

Color vision, a feature of visual perception, is an ability to perceive differences between light composed of different frequencies independently of light intensity.

The visual system is the physiological basis of visual perception. The system detects, transduces and interprets information concerning light within the visible range to construct an image and build a mental model of the surrounding environment. The visual system is associated with the eye and functionally divided into the optical system and the neural system.

A Brodmann area is a region of the cerebral cortex, in the human or other primate brain, defined by its cytoarchitecture, or histological structure and organization of cells. The concept was first introduced by the German anatomist Korbinian Brodmann in the early 20th century. Brodmann mapped the human brain based on the varied cellular structure across the cortex and identified 52 distinct regions, which he numbered 1 to 52. These regions, or Brodmann areas, correspond with diverse functions including sensation, motor control, and cognition.

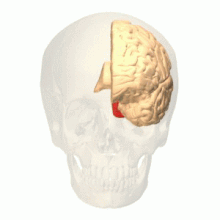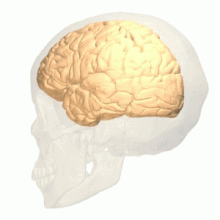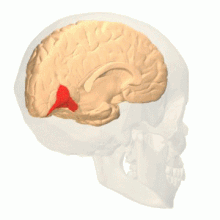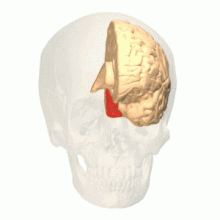
The occipital lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The name derives from its position at the back of the head, from the Latin ob, 'behind', and caput, 'head'.

The fusiform gyrus, also known as the lateral occipitotemporal gyrus,is part of the temporal lobe and occipital lobe in Brodmann area 37. The fusiform gyrus is located between the lingual gyrus and parahippocampal gyrus above, and the inferior temporal gyrus below. Though the functionality of the fusiform gyrus is not fully understood, it has been linked with various neural pathways related to recognition. Additionally, it has been linked to various neurological phenomena such as synesthesia, dyslexia, and prosopagnosia.
Visual processing is a term that is used to refer to the brain's ability to use and interpret visual information from the world around us. The process of converting light energy into a meaningful image is a complex process that is facilitated by numerous brain structures and higher level cognitive processes. On an anatomical level, light energy first enters the eye through the cornea, where the light is bent. After passing through the cornea, light passes through the pupil and then lens of the eye, where it is bent to a greater degree and focused upon the retina. The retina is where a group of light-sensing cells, called photoreceptors are located. There are two types of photoreceptors: rods and cones. Rods are sensitive to dim light and cones are better able to transduce bright light. Photoreceptors connect to bipolar cells, which induce action potentials in retinal ganglion cells. These retinal ganglion cells form a bundle at the optic disc, which is a part of the optic nerve. The two optic nerves from each eye meet at the optic chiasm, where nerve fibers from each nasal retina cross which results in the right half of each eye's visual field being represented in the left hemisphere and the left half of each eye's visual fields being represented in the right hemisphere. The optic tract then diverges into two visual pathways, the geniculostriate pathway and the tectopulvinar pathway, which send visual information to the visual cortex of the occipital lobe for higher level processing.

The parahippocampal gyrus is a grey matter cortical region of the brain that surrounds the hippocampus and is part of the limbic system. The region plays an important role in memory encoding and retrieval. It has been involved in some cases of hippocampal sclerosis. Asymmetry has been observed in schizophrenia.

Retinotopy is the mapping of visual input from the retina to neurons, particularly those neurons within the visual stream. For clarity, 'retinotopy' can be replaced with 'retinal mapping', and 'retinotopic' with 'retinally mapped'.

The inferior temporal gyrus is one of three gyri of the temporal lobe and is located below the middle temporal gyrus, connected behind with the inferior occipital gyrus; it also extends around the infero-lateral border on to the inferior surface of the temporal lobe, where it is limited by the inferior sulcus. This region is one of the higher levels of the ventral stream of visual processing, associated with the representation of objects, places, faces, and colors. It may also be involved in face perception, and in the recognition of numbers and words.

The lingual gyrus, also known as the medialoccipitotemporal gyrus, is a brain structure that is linked to processing vision, especially related to letters. It is thought to also play a role in analysis of logical conditions and encoding visual memories. It is named after its shape, which is somewhat similar to a tongue. Contrary to the name, the region has little to do with speech.

Cerebral achromatopsia is a type of color blindness caused by damage to the cerebral cortex of the brain, rather than abnormalities in the cells of the eye's retina. It is often confused with congenital achromatopsia but underlying physiological deficits of the disorders are completely distinct. A similar, but distinct, deficit called color agnosia exists in which a person has intact color perception but has deficits in color recognition, such as knowing which color they are looking at.

The fusiform face area is a part of the human visual system that is specialized for facial recognition. It is located in the inferior temporal cortex (IT), in the fusiform gyrus.
In cognitive neuroscience, visual modularity is an organizational concept concerning how vision works. The way in which the primate visual system operates is currently under intense scientific scrutiny. One dominant thesis is that different properties of the visual world require different computational solutions which are implemented in anatomically/functionally distinct regions that operate independently – that is, in a modular fashion.
Discrete categories of objects such as faces, body parts, tools, animals and buildings have been associated with preferential activation in specialised areas of the cerebral cortex, leading to the suggestion that they may be produced separately in discrete neural regions.
Globs are millimeter-sized color modules found beyond the visual area V2 in the brain's color processing ventral pathway. They are scattered throughout the posterior inferior temporal cortex in an area called the V4 complex. They are clustered by color preference, and organized as color columns. They are the first part of the brain in which color is processed in terms of the full range of hues found in color space.
Visual object recognition refers to the ability to identify the objects in view based on visual input. One important signature of visual object recognition is "object invariance", or the ability to identify objects across changes in the detailed context in which objects are viewed, including changes in illumination, object pose, and background context.
Binocular neurons are neurons in the visual system that assist in the creation of stereopsis from binocular disparity. They have been found in the primary visual cortex where the initial stage of binocular convergence begins. Binocular neurons receive inputs from both the right and left eyes and integrate the signals together to create a perception of depth.
The occipital face area (OFA) is a region of the human cerebral cortex which is specialised for face perception. The OFA is located on the lateral surface of the occipital lobe adjacent to the inferior occipital gyrus. The OFA comprises a network of brain regions including the fusiform face area (FFA) and posterior superior temporal sulcus (STS) which support facial processing.