A honey bee is a eusocial flying insect within the genus Apis of the bee clade, all native to mainland Afro-Eurasia. After bees spread naturally throughout Africa and Eurasia, humans became responsible for the current cosmopolitan distribution of honey bees, introducing multiple subspecies into South America, North America, and Australia.

The Africanized bee, also known as the Africanized honey bee (AHB) and colloquially as the "killer bee", is a hybrid of the western honey bee, produced originally by crossbreeding of the East African lowland honey bee (A. m. scutellata) with various European honey bee subspecies such as the Italian honey bee (A. m. ligustica) and the Iberian honey bee (A. m. iberiensis).

A queen bee is typically an adult, mated female (gyne) that lives in a colony or hive of honey bees. With fully developed reproductive organs, the queen is usually the mother of most, if not all, of the bees in the beehive. Queens are developed from larvae selected by worker bees and specially fed in order to become sexually mature. There is normally only one adult, mated queen in a hive, in which case the bees will usually follow and fiercely protect her.

Apis mellifera ligustica is the Italian bee or the Italian Honey bee which is a subspecies of the western honey bee.

Aethina tumida,commonly known as small hive beetle (SHB), is a beekeeping pest. It is native to sub-Saharan Africa, but has spread to many other regions, including North America, Australia, and the Philippines.

Bombus terrestris, the buff-tailed bumblebee or large earth bumblebee, is one of the most numerous bumblebee species in Europe. It is one of the main species used in greenhouse pollination, and so can be found in many countries and areas where it is not native, such as Tasmania. Moreover, it is a eusocial insect with an overlap of generations, a division of labour, and cooperative brood care. The queen is monogamous which means she mates with only one male. B. terrestris workers learn flower colours and forage efficiently.

The dwarf honey bee, Apis florea, is one of two species of small, wild honey bees of southern and southeastern Asia. It has a much wider distribution than its sister species, Apis andreniformis. First identified in the late 18th century, Apis florea is unique for its morphology, foraging behavior and defensive mechanisms like making a piping noise. Apis florea have open nests and small colonies, which makes them more susceptible to predation than cavity nesters with large numbers of defensive workers. These honey bees are important pollinators and therefore commodified in countries like Cambodia.

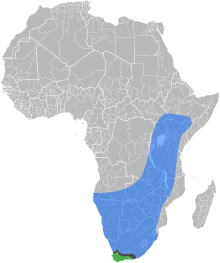
The Cape honey bee or Cape bee is a southern South African subspecies of the western honey bee. They play a major role in South African agriculture and the economy of the Western Cape by pollinating crops and producing honey in the Western Cape region of South Africa. The species is endemic to the Western Cape region of South Africa on the coastal side of the Cape Fold mountain range.

Apis andreniformis, or the black dwarf honey bee, is a relatively rare species of honey bee whose native habitat is the tropical and subtropical regions of Southeast Asia.

Apis koschevnikovi, Koschevnikov's honey bee, is a species of honey bee which inhabits Malaysian and Indonesian Borneo, where it lives sympatrically with other honey bee species such as Apis cerana.

Apis dorsata, the rock bee or giant honey bee, is a honey bee of South and Southeast Asia. They are typically around 17–20 mm (0.7–0.8 in) long and nests are mainly built in exposed places far off the ground, like on tree limbs, under cliff overhangs, and under buildings. These social bees are known for their aggressive defense strategies and vicious behavior when disturbed. Though not domesticated, indigenous peoples have traditionally used this species as a source of honey and beeswax, a practice known as honey hunting.
A tremble dance is a dance performed by forager honey bees of the species Apis mellifera to recruit more receiver honey bees to collect nectar from the workers.

The western honey bee or European honey bee is the most common of the 7–12 species of honey bees worldwide. The genus name Apis is Latin for "bee", and mellifera is the Latin for "honey-bearing" or "honey carrying", referring to the species' production of honey.

Eusociality is the highest level of organization of sociality. It is defined by the following characteristics: cooperative brood care, overlapping generations within a colony of adults, and a division of labor into reproductive and non-reproductive groups. The division of labor creates specialized behavioral groups within an animal society which are sometimes referred to as 'castes'. Eusociality is distinguished from all other social systems because individuals of at least one caste usually lose the ability to perform behaviors characteristic of individuals in another caste. Eusocial colonies can be viewed as superorganisms.

Apis cerana, the eastern honey bee, Asiatic honey bee or Asian honey bee, is a species of honey bee native to South, Southeast and East Asia. This species is the sister species of Apis koschevnikovi and both are in the same subgenus as the western (European) honey bee, Apis mellifera. A. cerana is known to live sympatrically along with Apis koschevnikovi within the same geographic location. Apis cerana colonies are known for building nests consisting of multiple combs in cavities containing a small entrance, presumably for defense against invasion by individuals of another nest. The diet of this honey bee species consists mostly of pollen and nectar, or honey. Moreover, Apis cerana is known for its highly social behavior, reflective of its classification as a type of honey bee.

Bumblebees, like the honeybee collect nectar and pollen from flowers and store them for food. Many individuals must be recruited to forage for food to provide for the hive. Some bee species have highly developed ways of communicating with each other about the location and quality of food resources ranging from physical to chemical displays.

Worker policing is a behavior seen in colonies of social hymenopterans whereby worker females eat or remove eggs that have been laid by other workers rather than those laid by a queen. Worker policing ensures that the offspring of the queen will predominate in the group. In certain species of bees, ants and wasps, workers or the queen may also act aggressively towards fertile workers. Worker policing has been suggested as a form of coercion to promote the evolution of altruistic behavior in eusocial insect societies.

Melipona beecheii is a species of eusocial stingless bee. It is native to Central America from the Yucatán Peninsula in the north to Costa Rica in the south. M. beecheii was cultivated in the Yucatán Peninsula starting in the pre-Columbian era by the ancient Maya civilization. The Mayan name for M. beecheii is xunan kab, which translates roughly to "regal lady bee". M. beecheii serves as the subject of various Mayan religious ceremonies.