
Mosasaurs are an extinct group of large aquatic reptiles within the family Mosasauridae that lived during the Late Cretaceous. Their first fossil remains were discovered in a limestone quarry at Maastricht on the Meuse in 1764. They belong to the order Squamata, which includes lizards and snakes.

Mosasaurus is the type genus of the mosasaurs, an extinct group of aquatic squamate reptiles. It lived from about 82 to 66 million years ago during the Campanian and Maastrichtian stages of the Late Cretaceous. The genus was one of the first Mesozoic marine reptiles known to science—the first fossils of Mosasaurus were found as skulls in a chalk quarry near the Dutch city of Maastricht in the late 18th century, and were initially thought to be crocodiles or whales. One skull discovered around 1780 was famously nicknamed the "great animal of Maastricht". In 1808, naturalist Georges Cuvier concluded that it belonged to a giant marine lizard with similarities to monitor lizards but otherwise unlike any known living animal. This concept was revolutionary at the time and helped support the then-developing ideas of extinction. Cuvier did not designate a scientific name for the animal; this was done by William Daniel Conybeare in 1822 when he named it Mosasaurus in reference to its origin in fossil deposits near the Meuse River. The exact affinities of Mosasaurus as a squamate remain controversial, and scientists continue to debate whether its closest living relatives are monitor lizards or snakes.
Tylosaurus is a genus of mosasaur, a large, predatory marine reptile closely related to modern monitor lizards and snakes, from the Late Cretaceous.

The Mosasaurinae are a subfamily of mosasaurs, a diverse group of Late Cretaceous marine squamates. Members of the subfamily are informally and collectively known as "mosasaurines" and their fossils have been recovered from every continent except for South America.

The Tylosaurinae are a subfamily of mosasaurs, a diverse group of Late Cretaceous marine squamates. Members of the subfamily are informally and collectively known as "tylosaurines" and have been recovered from every continent except for South America. The subfamily includes the genera Tylosaurus, Taniwhasaurus, and Kaikaifilu, although some scientists argue that only Tylosaurus and Taniwhasaurus should be included.

Globidens is an extinct genus of mosasaurid oceanic lizard classified as part of the Globidensini tribe in the Mosasaurinae subfamily.

Prognathodon is an extinct genus of marine lizard belonging to the mosasaur family. It is classified as part of the Mosasaurinae subfamily, alongside genera like Mosasaurus and Clidastes. Prognathodon has been recovered from deposits ranging in age from the Campanian to the Maastrichtian in the Middle East, Europe, New Zealand, and North America.
Clidastes is an extinct genus of marine lizard belonging to the mosasaur family. It is classified as part of the Mosasaurinae subfamily, alongside genera like Mosasaurus and Prognathodon. Clidastes is known from deposits ranging in age from the Coniacian to the early Campanian in the United States.
Carinodens is an extinct genus of Cretaceous marine lizard belonging to the mosasaur family. "Carinodens" means "keel teeth" and was named in 1969 as a replacement name for Compressidens, "compressed teeth", which was already in use for a gadilidan scaphopod mollusk.

Pluridens is an extinct genus of marine lizard belonging to the Mosasauridae. Pluridens is placed in the subfamily Halisaurinae with the genera Phosphorosaurus, Eonatator and Halisaurus. Compared to related halisaurines, Pluridens had longer jaws with more teeth, and smaller eyes. It also grew large size, measuring 5–6 m (16–20 ft) long and perhaps over 9 m (30 ft) in some individuals. The jaws in some specimens are robust, and sometimes show injuries suggestive of combat. The jaws may have been used for fighting over mates or territories.
Goronyosaurus is an extinct genus of marine lizard belonging to the mosasaur family. Fossils of Goronyosaurus are exclusively known from the Late Maastrichtian of the Iullemmeden Basin in West Africa, specifically the Dukamaje Formation of Niger and Nigeria and Farin Doutchi Formation of Niger. The type specimen was first described in 1930 as Mosasaurus nigeriensis, but subsequent remains revealed a highly unique set of adaptations that prompted the species to be reclassified as the only species of the new genus Goronyosaurus in 1972. These unique adaptations have made Goronyosaurus notoriously difficult to classify within the Mosasauridae and it is often left out of phylogenetic analyses, although most authors agree that Goronyosaurus belonged to Mosasauridae.

Taniwhasaurus is an extinct genus of mosasaurs that lived during the Campanian stage of the Late Cretaceous. It is a member of the subfamily Tylosaurinae, a lineage of mosasaurs characterized by a long toothless conical rostrum. Two valid species are attached to the genus, T. oweni and T. antarcticus, known respectively from the fossil record of present-day New Zealand and Antarctica. Two other species have been nominally classified within the genus, T. 'capensis' and T. 'mikasaensis', recorded in present-day South Africa and Japan, but their attribution remains problematic due to the fragmentary state of their fossils. The generic name literally means "taniwha lizard", referring to a supernatural aquatic creature from Māori mythology.

Liodon is a dubious genus of mosasaur from the Late Cretaceous, known from fragmentary fossils discovered in St James' Pit, England and possibly also the Ouled Abdoun Basin of Morocco. Though dubious and of uncertain phylogenetic affinities, Liodon was historically a highly important taxon in mosasaur systematics, being one of the genera on which the family Mosasauridae was based.

Plesiotylosaurus, meaning "near Tylosaurus", is an extinct genus of marine lizard belonging to the mosasaur family. It is classified as part of the Mosasaurinae subfamily, alongside genera like Mosasaurus and Prognathodon. The genus contains one species, Plesiotylosaurus crassidens, recovered from deposits of Middle Maastrichtian age in the Moreno Formation in California.

Tethysaurus is an extinct genus of tethysaurine mosasauroid from the Early Turonian period. The only species is Tethysaurus nopcsai.

Mosasaurini is an extinct tribe of mosasaurine mosasaurs who lived during the Late Cretaceous and whose fossils have been found in North America, South America, Europe, Africa and Oceania, with questionable occurrences in Asia. They are highly derived mosasaurs, containing genera like Plotosaurus, having unique adaptations to fast swimming speeds, or Mosasaurus, which is among the largest known marine reptiles.

This timeline of mosasaur research is a chronologically ordered list of important fossil discoveries, controversies of interpretation, and taxonomic revisions of mosasaurs, a group of giant marine lizards that lived during the Late Cretaceous Epoch. Although mosasaurs went extinct millions of years before humans evolved, humans have coexisted with mosasaur fossils for millennia. Before the development of paleontology as a formal science, these remains would have been interpreted through a mythological lens. Myths about warfare between serpentine water monsters and aerial thunderbirds told by the Native Americans of the modern western United States may have been influenced by observations of mosasaur fossils and their co-occurrence with creatures like Pteranodon and Hesperornis.

Kaikaifilu is an extinct genus of large mosasaurs that lived during the Late Cretaceous (Maastrichtian) in what is now northern Antarctica. The only species known, K. hervei, was described in 2017 from an incomplete specimen discovered in the López de Bertodano Formation, in Seymour Island, Antarctic Peninsula. The taxon is named in reference to Coi Coi-Vilu, a reptilian ocean deity of the Mapuche cosmology. Early observations of the holotype classify it as a member of the subfamily Tylosaurinae. However, later observations note that several characteristics show that this attribution is problematic.

Xenodens is an extinct genus of marine lizard belonging to the mosasaur family. It currently contains a single species, X. calminechari, which is known from Late Maastrichtian phosphate deposits in the Ouled Abdoun Basin, Morocco. Its closest known relative is believed to be the durophagous Carinodens.
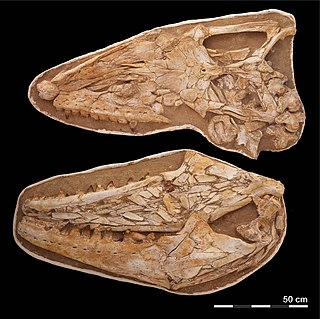
Thalassotitan is an extinct genus of large mosasaurs that lived during the late Maastrichtian of the Cretaceous period in what is now Morocco, around 66 million years ago. The only known species is T. atrox, described in 2022 from fossils discovered in the Ouled Abdoun Basin, where many other mosasaurs have been found. It was assigned to the tribe Prognathodontini alongside other mosasaurs like Prognathodon and Gnathomortis. The prognathodontines are separated from other mosasaurs based on their massive jaws and robust teeth.





