
Perception is the organization, identification, and interpretation of sensory information in order to represent and understand the presented information or environment. All perception involves signals that go through the nervous system, which in turn result from physical or chemical stimulation of the sensory system. Vision involves light striking the retina of the eye; smell is mediated by odor molecules; and hearing involves pressure waves.

The visual cortex of the brain is the area of the cerebral cortex that processes visual information. It is located in the occipital lobe. Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex. The area of the visual cortex that receives the sensory input from the lateral geniculate nucleus is the primary visual cortex, also known as visual area 1 (V1), Brodmann area 17, or the striate cortex. The extrastriate areas consist of visual areas 2, 3, 4, and 5.
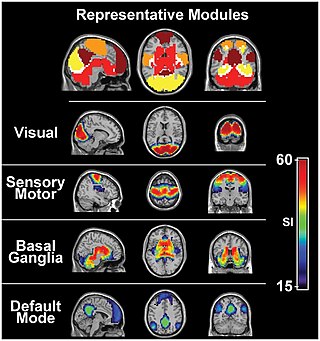
The sensory nervous system is a part of the nervous system responsible for processing sensory information. A sensory system consists of sensory neurons, neural pathways, and parts of the brain involved in sensory perception and interoception. Commonly recognized sensory systems are those for vision, hearing, touch, taste, smell, balance and visceral sensation. Sense organs are transducers that convert data from the outer physical world to the realm of the mind where people interpret the information, creating their perception of the world around them.
The receptive field, or sensory space, is a delimited medium where some physiological stimuli can evoke a sensory neuronal response in specific organisms.

Motion perception is the process of inferring the speed and direction of elements in a scene based on visual, vestibular and proprioceptive inputs. Although this process appears straightforward to most observers, it has proven to be a difficult problem from a computational perspective, and difficult to explain in terms of neural processing.
Multisensory integration, also known as multimodal integration, is the study of how information from the different sensory modalities may be integrated by the nervous system. A coherent representation of objects combining modalities enables animals to have meaningful perceptual experiences. Indeed, multisensory integration is central to adaptive behavior because it allows animals to perceive a world of coherent perceptual entities. Multisensory integration also deals with how different sensory modalities interact with one another and alter each other's processing.

Illusory contours or subjective contours are visual illusions that evoke the perception of an edge without a luminance or color change across that edge. Illusory brightness and depth ordering often accompany illusory contours. Friedrich Schumann is often credited with the discovery of illusory contours around the beginning of the 20th century, but they are present in art dating to the Middle Ages. Gaetano Kanizsa’s 1976 Scientific American paper marked the resurgence of interest in illusory contours for vision scientists.

The Chubb illusion is an optical illusion or error in visual perception in which the apparent contrast of an object varies substantially to most viewers depending on its relative contrast to the field on which it is displayed. These visual illusions are of particular interest to researchers because they may provide valuable insights in regard to the workings of human visual systems.
Repetition priming refers to improvements in a behavioural response when stimuli are repeatedly presented. The improvements can be measured in terms of accuracy or reaction time and can occur when the repeated stimuli are either identical or similar to previous stimuli. These improvements have been shown to be cumulative, so as the number of repetitions increases the responses get continually faster up to a maximum of around seven repetitions. These improvements are also found when the repeated items are changed slightly in terms of orientation, size and position. The size of the effect is also modulated by the length of time the item is presented for and the length time between the first and subsequent presentations of the repeated items.
Flash suppression is a phenomenon of visual perception in which an image presented to one eye is suppressed by a flash of another image presented to the other eye.

Motion Induced Blindness (MIB), also known as Bonneh's illusion is a visual illusion in which a large, continuously moving pattern erases from perception some small, continuously presented, stationary dots when one looks steadily at the center of the display. It was discovered by Bonneh, Cooperman, and Sagi (2001), who used a swarm of blue dots moving on a virtual sphere as the larger pattern and three small yellow dots as the smaller pattern. They found that after about 10 seconds, one or more of the dots disappeared for brief, random times.

The neural correlates of consciousness (NCC) are the minimal set of neuronal events and mechanisms sufficient for the occurrence of the mental states to which they are related. Neuroscientists use empirical approaches to discover neural correlates of subjective phenomena; that is, neural changes which necessarily and regularly correlate with a specific experience. The set should be minimal because, under the materialist assumption that the brain is sufficient to give rise to any given conscious experience, the question is which of its components are necessary to produce it.
Feature detection is a process by which the nervous system sorts or filters complex natural stimuli in order to extract behaviorally relevant cues that have a high probability of being associated with important objects or organisms in their environment, as opposed to irrelevant background or noise.
Transsaccadic memory is the neural process that allows humans to perceive their surroundings as a seamless, unified image despite rapid changes in fixation points. Transsaccadic memory is a relatively new topic of interest in the field of psychology. Conflicting views and theories have spurred several types of experiments intended to explain transsaccadic memory and the neural mechanisms involved.

A phantom contour is a type of illusory contour. Most illusory contours are seen in still images, such as the Kanizsa triangle and the Ehrenstein illusion. A phantom contour, however, is perceived in the presence of moving or flickering images with contrast reversal. The rapid, continuous alternation between opposing, but correlated, adjacent images creates the perception of a contour that is not physically present in the still images. Quaid et al. have also authored a PhD thesis on the phantom contour illusion and its spatiotemporal limits which maps out limits and proposes mechanisms for its perception centering around magnocellularly driven visual area MT.

Due to the effect of a spatial context or temporal context, the perceived orientation of a test line or grating pattern can appear tilted away from its physical orientation. The tilt illusion (TI) is the phenomenon that the perceived orientation of a test line or grating is altered by the presence of surrounding lines or grating with a different orientation. And the tilt aftereffect (TAE) is the phenomenon that the perceived orientation is changed after prolonged inspection of another oriented line or grating.
Biased competition theory advocates the idea that each object in the visual field competes for cortical representation and cognitive processing. This theory suggests that the process of visual processing can be biased by other mental processes such as bottom-up and top-down systems which prioritize certain features of an object or whole items for attention and further processing. Biased competition theory is, simply stated, the competition of objects for processing. This competition can be biased, often toward the object that is currently attended in the visual field, or alternatively toward the object most relevant to behavior.
Binocular neurons are neurons in the visual system that assist in the creation of stereopsis from binocular disparity. They have been found in the primary visual cortex where the initial stage of binocular convergence begins. Binocular neurons receive inputs from both the right and left eyes and integrate the signals together to create a perception of depth.
Surround suppression is where the relative firing rate of a neuron may under certain conditions decrease when a particular stimulus is enlarged. It has been observed in electrophysiology studies of the brain and has been noted in many sensory neurons, most notably in the early visual system. Surround suppression is defined as a reduction in the activity of a neuron in response to a stimulus outside its classical receptive field.

Russell L. De Valois was an American scientist recognized for his pioneering research on spatial and color vision.
