Related Research Articles

Color blindness or color vision deficiency (CVD) is the decreased ability to see color or differences in color. The severity of color blindness ranges from mostly unnoticeable to full absence of color perception. Color blindness is usually an inherited problem or variation in the functionality of one or more of the three classes of cone cells in the retina, which mediate color vision. The most common form is caused by a genetic condition called congenital red–green color blindness, which affects up to 1 in 12 males (8%) and 1 in 200 females (0.5%). The condition is more prevalent in males, because the opsin genes responsible are located on the X chromosome. Rarer genetic conditions causing color blindness include congenital blue–yellow color blindness, blue cone monochromacy, and achromatopsia. Color blindness can also result from physical or chemical damage to the eye, the optic nerve, parts of the brain, or from medication toxicity. Color vision also naturally degrades in old age.

The retina is the innermost, light-sensitive layer of tissue of the eye of most vertebrates and some molluscs. The optics of the eye create a focused two-dimensional image of the visual world on the retina, which then processes that image within the retina and sends nerve impulses along the optic nerve to the visual cortex to create visual perception. The retina serves a function which is in many ways analogous to that of the film or image sensor in a camera.
Achromatopsia, also known as Rod monochromacy, is a medical syndrome that exhibits symptoms relating to five conditions, most notably monochromacy. Historically, the name referred to monochromacy in general, but now typically refers only to an autosomal recessive congenital color vision condition. The term is also used to describe cerebral achromatopsia, though monochromacy is usually the only common symptom. The conditions include: monochromatic color blindness, poor visual acuity, and day-blindness. The syndrome is also present in an incomplete form that exhibits milder symptoms, including residual color vision. Achromatopsia is estimated to affect 1 in 30,000 live births worldwide.

Color vision, a feature of visual perception, is an ability to perceive differences between light composed of different frequencies independently of light intensity.

Tetrachromacy is the condition of possessing four independent channels for conveying color information, or possessing four types of cone cell in the eye. Organisms with tetrachromacy are called tetrachromats.

Trichromacy or trichromatism is the possession of three independent channels for conveying color information, derived from the three different types of cone cells in the eye. Organisms with trichromacy are called trichromats.
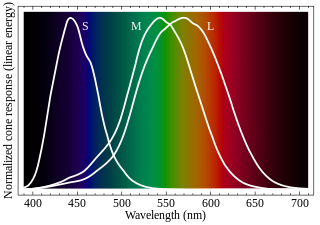
Cone cells or cones are photoreceptor cells in the retinas of vertebrates' eyes. They respond differently to light of different wavelengths, and the combination of their responses is responsible for color vision. Cones function best in relatively bright light, called the photopic region, as opposed to rod cells, which work better in dim light, or the scotopic region. Cone cells are densely packed in the fovea centralis, a 0.3 mm diameter rod-free area with very thin, densely packed cones which quickly reduce in number towards the periphery of the retina. Conversely, they are absent from the optic disc, contributing to the blind spot. There are about six to seven million cones in a human eye, with the highest concentration being towards the macula.
Dichromacy is the state of having two types of functioning photoreceptors, called cone cells, in the eyes. Organisms with dichromacy are called dichromats. Dichromats require only two primary colors to be able to represent their visible gamut. By comparison, trichromats need three primary colors, and tetrachromats need four. Likewise, every color in a dichromat's gamut can be evoked with monochromatic light. By comparison, every color in a trichromat's gamut can be evoked with a combination of monochromatic light and white light.

Monochromacy is the ability of organisms to perceive only light intensity without respect to spectral composition. Organisms with monochromacy lack color vision and can only see in shades of grey ranging from black to white. Organisms with monochromacy are called monochromats. Many mammals, such as cetaceans, the owl monkey and the Australian sea lion are monochromats. In humans, monochromacy is one among several other symptoms of severe inherited or acquired diseases, including achromatopsia or blue cone monochromacy, together affecting about 1 in 30,000 people.

Melanopsin is a type of photopigment belonging to a larger family of light-sensitive retinal proteins called opsins and encoded by the gene Opn4. In the mammalian retina, there are two additional categories of opsins, both involved in the formation of visual images: rhodopsin and photopsin in the rod and cone photoreceptor cells, respectively.

Blue-sensitive opsin is a protein that in humans is encoded by the OPN1SW gene.

Green-sensitive opsin is a protein that in humans is encoded by the OPN1MW gene. OPN1MW2 is a similar opsin.
OPN1LW is a gene on the X chromosome that encodes for long wave sensitive (LWS) opsin, or red cone photopigment. It is responsible for perception of visible light in the yellow-green range on the visible spectrum. The gene contains 6 exons with variability that induces shifts in the spectral range. OPN1LW is subject to homologous recombination with OPN1MW, as the two have very similar sequences. These recombinations can lead to various vision problems, such as red-green colourblindness and blue monochromacy. The protein encoded is a G-protein coupled receptor with embedded 11-cis-retinal, whose light excitation causes a cis-trans conformational change that begins the process of chemical signalling to the brain.

The evolution of color vision in primates is highly unusual compared to most eutherian mammals. A remote vertebrate ancestor of primates possessed tetrachromacy, but nocturnal, warm-blooded, mammalian ancestors lost two of four cones in the retina at the time of dinosaurs. Most teleost fish, reptiles and birds are therefore tetrachromatic while most mammals are strictly dichromats, the exceptions being some primates and marsupials, who are trichromats, and many marine mammals, who are monochromats.
Color vision, a proximate adaptation of the vision sensory modality, allows for the discrimination of light based on its wavelength components.
Gene therapy using lentiviral vectors was being explored in early stage trials as of 2009.
Retinal gene therapy holds a promise in treating different forms of non-inherited and inherited blindness.
Blue cone monochromacy (BCM) is an inherited eye disease that causes severe color blindness, poor visual acuity, nystagmus and photophobia due to the absence of functional red (L) and green (M) cone photoreceptor cells in the retina. BCM is a recessive X-linked disease and almost exclusively affects XY karyotypes.

Color blind glasses or color correcting lenses are light filters, usually in the form of glasses or contact lenses, that attempt to alleviate color blindness, by bringing deficient color vision closer to normal color vision or to make certain color tasks easier to accomplish. Despite its viral status, the academic literature is generally skeptical of the efficacy of color correcting lenses.
Congenital red–green color blindness is an inherited condition that is the root cause of the majority of cases of color blindness. It has no significant symptoms aside from its minor to moderate effect on color vision. It is caused by variation in the functionality of the red and/or green opsin proteins, which are the photosensitive pigment in the cone cells of the retina, which mediate color vision. Males are more likely to inherit red–green color blindness than females, because the genes for the relevant opsins are on the X chromosome. Screening for congenital red–green color blindness is typically performed with the Ishihara or similar color vision test. There is no cure for color blindness.
References
- 1 2 3 4 Kandel ER, Schwartz JH, Jessel TM, eds. (2000). "Ch. 29: Color Vision". Principles of Neural Science . McGraw-Hill Professional. ISBN 978-0-8385-7701-1.
- ↑ Bennicelli J, Wright JF, Komaromy A, Jacobs JB, Hauck B, Zelenaia O, et al. (March 2008). "Reversal of blindness in animal models of leber congenital amaurosis using optimized AAV2-mediated gene transfer". Molecular Therapy. 16 (3): 458–65. doi:10.1038/sj.mt.6300389. PMC 2842085 . PMID 18209734.
- ↑ Jacobson SG, Boye SL, Aleman TS, Conlon TJ, Zeiss CJ, Roman AJ, et al. (August 2006). "Safety in nonhuman primates of ocular AAV2-RPE65, a candidate treatment for blindness in Leber congenital amaurosis". Human Gene Therapy. 17 (8): 845–58. doi:10.1089/hum.2006.17.845. PMID 16942444.
- ↑ Jacobs GH, Williams GA, Cahill H, Nathans J (March 2007). "Emergence of novel color vision in mice engineered to express a human cone photopigment". Science. 315 (5819): 1723–5. Bibcode:2007Sci...315.1723J. doi:10.1126/science.1138838. PMID 17379811. S2CID 85273369.
- ↑ Mauck MC, Mancuso K, Kuchenbecker JA, Connor TB, Hauswirth WW, Neitz J, Neitz M (2008). "Longitudinal evaluation of expression of virally delivered transgenes in gerbil cone photoreceptors". Visual Neuroscience. 25 (3): 273–82. doi:10.1017/S0952523808080577. PMC 2643299 . PMID 18598398.
- 1 2 3 4 5 6 7 Mancuso K, Hauswirth WW, Li Q, Connor TB, Kuchenbecker JA, Mauck MC, et al. (October 2009). "Gene therapy for red-green colour blindness in adult primates". Nature. 461 (7265): 784–7. Bibcode:2009Natur.461..784M. doi:10.1038/nature08401. PMC 2782927 . PMID 19759534.
- ↑ Alexander JJ, Umino Y, Everhart D, Chang B, Min SH, Li Q, et al. (June 2007). "Restoration of cone vision in a mouse model of achromatopsia". Nature Medicine. 13 (6): 685–7. doi:10.1038/nm1596. PMC 3985124 . PMID 17515894.
- ↑ Komáromy AM, Alexander JJ, Rowlan JS, Garcia MM, Chiodo VA, Kaya A, et al. (July 2010). "Gene therapy rescues cone function in congenital achromatopsia". Human Molecular Genetics. 19 (13): 2581–93. doi:10.1093/hmg/ddq136. PMC 2883338 . PMID 20378608.
- ↑ Farahbakhsh M, Anderson EJ, Maimon-Mor RO, Rider A, Greenwood JA, Hirji N, et al. (August 2022). "A demonstration of cone function plasticity after gene therapy in achromatopsia". Brain. 145 (11): 3803–3815. doi:10.1093/brain/awac226. PMC 9679164 . PMID 35998912.
- ↑ McKyton, Ayelet; Marks Ohana, Devora; Nahmany, Einav; Banin, Eyal; Levin, Netta (July 2023). "Seeing color following gene augmentation therapy in achromatopsia". Current Biology. 33 (16): 3489–3494.e2. Bibcode:2023CBio...33E3489M. doi:10.1016/j.cub.2023.06.041. PMID 37433300. S2CID 259504295.
- ↑ Jackson, Justin; Xpress, Medical. "Gene therapy to restore color vision in complete achromatopsia patients shows modest improvement". medicalxpress.com. Retrieved 2023-08-28.
- 1 2 Giarmarco MM, Barborek R, Rowlan J, Rezeanu D, Bembry B, Neitz J, Neitz M, Kuchenbecker JA (June 2022). "Efficiency of gene therapy via intravitreal injection in primate cones". Investigative Ophthalmology & Visual Science.
- 1 2 Bennett J (December 2009). "Gene therapy for color blindness". The New England Journal of Medicine. 361 (25): 2483–4. doi:10.1056/NEJMcibr0908643. PMID 20018970.
- ↑ Grieger JC, Samulski RJ (2005). "Adeno-associated Virus as a Gene Therapy Vector: Vector Development, Production and Clinical Applications". Gene Therapy and Gene Delivery Systems. Advances in Biochemical Engineering/Biotechnology. Vol. 99. pp. 119–45. doi:10.1007/10_005. ISBN 3-540-28404-4. PMID 16568890.
- ↑ Hubel DH, Wiesel TN, LeVay S (April 1977). "Plasticity of ocular dominance columns in monkey striate cortex". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 278 (961): 377–409. Bibcode:1977RSPTB.278..377H. doi: 10.1098/rstb.1977.0050 . PMID 19791.
- ↑ Morgan MJ, Adam A, Mollon JD (June 1992). "Dichromats detect colour-camouflaged objects that are not detected by trichromats". Proceedings. Biological Sciences. 248 (1323): 291–5. Bibcode:1992RSPSB.248..291M. doi:10.1098/rspb.1992.0074. PMID 1354367. S2CID 35694740.