Production of offspring with combinations of traits that differ from those found in either parent
A model of meiotic recombination, initiated by a double-strand break or gap, followed by pairing with an homologous chromosome and strand invasion to initiate the recombinational repair process. Repair of the gap can lead to crossover (CO) or non-crossover (NCO) of the flanking regions. CO recombination is thought to occur by the Double Holliday Junction (DHJ) model, illustrated on the right. NCO recombinants are thought to occur primarily by the Synthesis Dependent Strand Annealing (SDSA) model, illustrated on the left. Most recombination events appear to be the SDSA type.
Genetic recombination (also known as genetic reshuffling) is the exchange of genetic material between different organisms which leads to production of offspring with combinations of traits that differ from those found in either parent. In eukaryotes, genetic recombination during meiosis can lead to a novel set of genetic information that can be further passed on from parents to offspring. Most recombination occurs naturally and can be classified into two types: (1) interchromosomal recombination, occurring through independent assortment of alleles whose loci are on different but homologous chromosomes (random orientation of pairs of homologous chromosomes in meiosis I); & (2) intrachromosomal recombination, occurring through crossing over.[1]
During meiosis in eukaryotes, genetic recombination involves the pairing of homologous chromosomes. This may be followed by information transfer between the chromosomes. The information transfer may occur without physical exchange (a section of genetic material is copied from one chromosome to another, without the donating chromosome being changed) (see SDSA – Synthesis Dependent Strand Annealing pathway in Figure); or by the breaking and rejoining of DNA strands, which forms new molecules of DNA (see DHJ pathway in Figure).
Recombination may also occur during mitosis in eukaryotes where it ordinarily involves the two sister chromosomes formed after chromosomal replication. In this case, new combinations of alleles are not produced since the sister chromosomes are usually identical. In meiosis and mitosis, recombination occurs between similar molecules of DNA (homologous sequences). In meiosis, non-sister homologous chromosomes pair with each other so that recombination characteristically occurs between non-sister homologues. In both meiotic and mitotic cells, recombination between homologous chromosomes is a common mechanism used in DNA repair.
Gene conversion – the process during which homologous sequences are made identical also falls under genetic recombination.
During meiosis, synapsis (the pairing of homologous chromosomes) ordinarily precedes genetic recombination.
Mechanism
Genetic recombination is catalyzed by many different enzymes. Recombinases are key enzymes that catalyse the strand transfer step during recombination. RecA, the chief recombinase found in Escherichia coli, is responsible for the repair of DNA double strand breaks (DSBs). In yeast and other eukaryotic organisms there are two recombinases required for repairing DSBs. The RAD51 protein is required for mitotic and meiotic recombination, whereas the DNA repair protein, DMC1, is specific to meiotic recombination. In the archaea, the ortholog of the bacterial RecA protein is RadA.
In bacteria there is regular genetic recombination, as well as ineffective transfer of genetic material, expressed as unsuccessful transfer or abortive transfer, which is any bacterial DNA transfer of the donor cell to recipients which have set the incoming DNA as part of the genetic material of the recipient.[citation needed][clarification needed] Abortive transfer was registered in the following transduction and conjugation.[clarification needed] In all cases, the transmitted fragment is diluted by the culture growth.[2][3][4]
In eukaryotes, recombination during meiosis is facilitated by chromosomal crossover. The crossover process leads to offspring having different combinations of genes from those of their parents, and can occasionally produce new chimericalleles.[citation needed] The shuffling of genes brought about by genetic recombination produces increased genetic variation. It also allows sexually reproducing organisms to avoid Muller's ratchet, in which the genomes of an asexualpopulation tend to accumulate more deleterious mutations over time than beneficial or reversing mutations.[citation needed]
Chromosomal crossover involves recombination between the paired chromosomes inherited from each of one's parents, generally occurring during meiosis.[citation needed] During prophase I (pachytene stage) the four available chromatids are in tight formation with one another.[citation needed] While in this formation, homologous sites on two chromatids can closely pair with one another, and may exchange genetic information.[5]
Because there is a small probability of recombination at any location along a chromosome, the frequency of recombination between two locations depends on the distance separating them.[citation needed] Therefore, for genes sufficiently distant on the same chromosome, the amount of crossover is high enough to destroy the correlation between alleles.[citation needed]
Tracking the movement of genes resulting from crossovers has proven quite useful to geneticists. Because two genes that are close together are less likely to become separated than genes that are farther apart, geneticists can deduce roughly how far apart two genes are on a chromosome if they know the frequency of the crossovers.[citation needed] Geneticists can also use this method to infer the presence of certain genes. Genes that typically stay together during recombination are said to be linked. One gene in a linked pair can sometimes be used as a marker to deduce the presence of the other gene. This is typically used to detect the presence of a disease-causing gene.[6]
The recombination frequency between two loci observed is the crossing-over value. It is the frequency of crossing over between two linked gene loci (markers), and depends on the distance between the genetic loci observed. For any fixed set of genetic and environmental conditions, recombination in a particular region of a linkage structure (chromosome) tends to be constant, and the same is then true for the crossing-over value which is used in the production of genetic maps.[2][7]
In gene conversion, a section of genetic material is copied from one chromosome to another, without the donating chromosome being changed. Gene conversion occurs at high frequency at the actual site of the recombination event during meiosis. It is a process by which a DNA sequence is copied from one DNA helix (which remains unchanged) to another DNA helix, whose sequence is altered. Gene conversion has often been studied in fungal crosses[8] where the 4 products of individual meioses can be conveniently observed. Gene conversion events can be distinguished as deviations in an individual meiosis from the normal 2:2 segregation pattern (e.g. a 3:1 pattern).
Nonhomologous recombination
Recombination can occur between DNA sequences that contain no sequence homology. This can cause chromosomal translocations, sometimes leading to cancer.
In genetic engineering, recombination can also refer to artificial and deliberate recombination of disparate pieces of DNA, often from different organisms, creating what is called recombinant DNA. A prime example of such a use of genetic recombination is gene targeting, which can be used to add, delete or otherwise change an organism's genes. This technique is important to biomedical researchers as it allows them to study the effects of specific genes. Techniques based on genetic recombination are also applied in protein engineering to develop new proteins of biological interest.
DNA damages caused by a variety of exogenous agents (e.g. UV light, X-rays, chemical cross-linking agents) can be repaired by homologous recombinational repair (HRR).[9][10] These findings suggest that DNA damages arising from natural processes, such as exposure to reactive oxygen species that are byproducts of normal metabolism, are also repaired by HRR. In humans, deficiencies in the gene products necessary for HRR during meiosis likely cause infertility[11] In humans, deficiencies in gene products necessary for HRR, such as BRCA1 and BRCA2, increase the risk of cancer (see DNA repair-deficiency disorder).
In bacteria, transformation is a process of gene transfer that ordinarily occurs between individual cells of the same bacterial species. Transformation involves integration of donor DNA into the recipient chromosome by recombination. This process appears to be an adaptation for repairing DNA damages in the recipient chromosome by HRR.[12] Transformation may provide a benefit to pathogenic bacteria by allowing repair of DNA damage, particularly damages that occur in the inflammatory, oxidizing environment associated with infection of a host.
When two or more viruses, each containing lethal genomic damages, infect the same host cell, the virus genomes can often pair with each other and undergo HRR to produce viable progeny. This process, referred to as multiplicity reactivation, has been studied in lambda and T4 bacteriophages,[13] as well as in several pathogenic viruses. In the case of pathogenic viruses, multiplicity reactivation may be an adaptive benefit to the virus since it allows the repair of DNA damages caused by exposure to the oxidizing environment produced during host infection.[12] See also reassortment.
Meiotic recombination
A molecular model for the mechanism of meiotic recombination presented by Anderson and Sekelsky[14] is outlined in the first figure in this article. Two of the four chromatids present early in meiosis (prophase I) are paired with each other and able to interact. Recombination, in this model, is initiated by a double-strand break (or gap) shown in the DNA molecule (chromatid) at the top of the figure. Other types of DNA damage may also initiate recombination. For instance, an inter-strand cross-link (caused by exposure to a cross-linking agent such as mitomycin C) can be repaired by HRR.
Two types of recombinant product are produced. Indicated on the right side is a "crossover" (CO) type, where the flanking regions of the chromosomes are exchanged, and on the left side, a "non-crossover" (NCO) type where the flanking regions are not exchanged. The CO type of recombination involves the intermediate formation of two "Holliday junctions" indicated in the lower right of the figure by two X-shaped structures in each of which there is an exchange of single strands between the two participating chromatids. This pathway is labeled in the figure as the DHJ (double-Holliday junction) pathway.
The NCO recombinants (illustrated on the left in the figure) are produced by a process referred to as "synthesis dependent strand annealing" (SDSA). Recombination events of the NCO/SDSA type appear to be more common than the CO/DHJ type.[15] The NCO/SDSA pathway contributes little to genetic variation, since the arms of the chromosomes flanking the recombination event remain in the parental configuration. Thus, explanations for the adaptive function of meiosis that focus exclusively on crossing-over are inadequate to explain the majority of recombination events.
Achiasmy and heterochiasmy
Achiasmy is the phenomenon where autosomal recombination is completely absent in one sex of a species. Achiasmatic chromosomal segregation is well documented in male Drosophila melanogaster. Heterochiasmy occurs when recombination rates differ between the sexes of a species.[16] This sexual dimorphic pattern in recombination rate has been observed in many species. In mammals, females most often have higher rates of recombination. The "Haldane-Huxley rule" states that achiasmy usually occurs in the heterogametic sex.[16]
RNA virus recombination
Numerous RNA viruses are capable of genetic recombination when at least two viral genomes are present in the same host cell.[17][18] Recombination is largely responsible for RNA virus diversity and immune evasion.[19] RNA recombination appears to be a major driving force in determining genome architecture and the course of viral evolution among picornaviridae ((+)ssRNA) (e.g. poliovirus).[20] In the retroviridae ((+)ssRNA)(e.g. HIV), damage in the RNA genome appears to be avoided during reverse transcription by strand switching, a form of recombination.[21][22]
Recombination in RNA viruses appears to be an adaptation for coping with genome damage.[17] Switching between template strands during genome replication, referred to as copy-choice recombination, was originally proposed to explain the positive correlation of recombination events over short distances in organisms with a DNA genome (see first Figure, SDSA pathway).[25]
Recombination can occur infrequently between animal viruses of the same species but of divergent lineages. The resulting recombinant viruses may sometimes cause an outbreak of infection in humans.[23]
Especially in coronaviruses, recombination may also occur even among distantly related evolutionary groups (subgenera), due to their characteristic transcription mechanism, that involves subgenomic mRNAs that are formed by template switching.[26][24]
When replicating its (+)ssRNA genome, the poliovirusRNA-dependent RNA polymerase (RdRp) is able to carry out recombination. Recombination appears to occur by a copy choice mechanism in which the RdRp switches (+)ssRNA templates during negative strand synthesis.[27] Recombination by RdRp strand switching also occurs in the (+)ssRNA plant carmoviruses and tombusviruses.[28]
Recombination appears to be a major driving force in determining genetic variability within coronaviruses, as well as the ability of coronavirus species to jump from one host to another and, infrequently, for the emergence of novel species, although the mechanism of recombination in is unclear.[23]
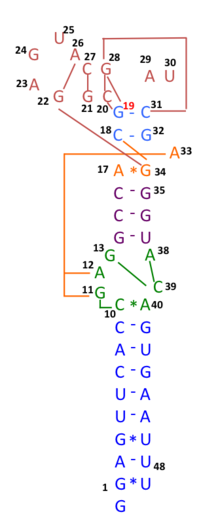
In early 2020, many genomic sequences of Australian SARS‐CoV‐2 isolates have deletions or mutations (29742G>A or 29742G>U; "G19A" or "G19U") in the s2m, suggesting that RNA recombination may have occurred in this RNA element. 29742G("G19"), 29744G("G21"), and 29751G("G28") were predicted as recombination hotspots.[29] During the first months of the COVID-19 pandemic, such a recombination event was suggested to have been a critical step in the evolution of SARS-CoV-2's ability to infect humans.[30]Linkage disequilibrium analysis confirmed that RNA recombination with the 11083G > T mutation also contributed to the increase of mutations among the viral progeny. The findings indicate that the 11083G > T mutation of SARS-CoV-2 spread during Diamond Princess shipboard quarantine and arose through de novo RNA recombination under positive selection pressure. In three patients on the Diamond Princess cruise, two mutations, 29736G > T and 29751G > T (G13 and G28) were located in Coronavirus 3′ stem-loop II-like motif (s2m) of SARS-CoV-2. Although s2m is considered an RNA motif highly conserved in 3' untranslated region among many coronavirus species, this result also suggests that s2m of SARS-CoV-2 is RNA recombination/mutation hotspot.[31]
Schematic representation of the s2m RNA secondary structure, with tertiary structural interactions indicated as long range contacts.
SARS-CoV-2's entire receptor binding motif appeared, based on preliminary observations, to have been introduced through recombination from coronaviruses of pangolins.[32] However, more comprehensive analyses later refuted this suggestion and showed that SARS-CoV-2 likely evolved solely within bats and with little or no recombination.[33][34]
Role of recombination in the origin of life
Nowak and Ohtsuki[35] noted that the origin of life (abiogenesis) is also the origin of biological evolution. They pointed out that all known life on earth is based on biopolymers and proposed that any theory for the origin of life must involve biological polymers that act as information carriers and catalysts. Lehman[36] argued that recombination was an evolutionary development as ancient as the origins of life. Smail et al.[37] proposed that in the primordial Earth, recombination played a key role in the expansion of the initially short informational polymers (presumed to be RNA) that were the precursors to life.
Meiosis is a special type of cell division of germ cells and apicomplexans in sexually-reproducing organisms that produces the gametes, the sperm or egg cells. It involves two rounds of division that ultimately result in four cells, each with only one copy of each chromosome (haploid). Additionally, prior to the division, genetic material from the paternal and maternal copies of each chromosome is crossed over, creating new combinations of code on each chromosome. Later on, during fertilisation, the haploid cells produced by meiosis from a male and a female will fuse to create a zygote, a cell with two copies of each chromosome again.
Chromosomal crossover, or crossing over, is the exchange of genetic material during sexual reproduction between two homologous chromosomes' non-sister chromatids that results in recombinant chromosomes. It is one of the final phases of genetic recombination, which occurs in the pachytene stage of prophase I of meiosis during a process called synapsis. Synapsis begins before the synaptonemal complex develops and is not completed until near the end of prophase I. Crossover usually occurs when matching regions on matching chromosomes break and then reconnect to the other chromosome.
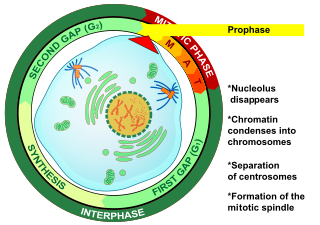
Prophase is the first stage of cell division in both mitosis and meiosis. Beginning after interphase, DNA has already been replicated when the cell enters prophase. The main occurrences in prophase are the condensation of the chromatin reticulum and the disappearance of the nucleolus.
A pair of homologous chromosomes, or homologs, are a set of one maternal and one paternal chromosome that pair up with each other inside a cell during fertilization. Homologs have the same genes in the same loci, where they provide points along each chromosome that enable a pair of chromosomes to align correctly with each other before separating during meiosis. This is the basis for Mendelian inheritance, which characterizes inheritance patterns of genetic material from an organism to its offspring parent developmental cell at the given time and area.
Sexual reproduction is an adaptive feature which is common to almost all multicellular organisms and various unicellular organisms. Currently, the adaptive advantage of sexual reproduction is widely regarded as a major unsolved problem in biology. As discussed below, one prominent theory is that sex evolved as an efficient mechanism for producing variation, and this had the advantage of enabling organisms to adapt to changing environments. Another prominent theory, also discussed below, is that a primary advantage of outcrossing sex is the masking of the expression of deleterious mutations. Additional theories concerning the adaptive advantage of sex are also discussed below. Sex does, however, come with a cost. In reproducing asexually, no time nor energy needs to be expended in choosing a mate and, if the environment has not changed, then there may be little reason for variation, as the organism may already be well-adapted. However, very few environments have not changed over the millions of years that reproduction has existed. Hence it is easy to imagine that being able to adapt to changing environment imparts a benefit. Sex also halves the amount of offspring a given population is able to produce. Sex, however, has evolved as the most prolific means of species branching into the tree of life. Diversification into the phylogenetic tree happens much more rapidly via sexual reproduction than it does by way of asexual reproduction.
RecQ helicase is a family of helicase enzymes initially found in Escherichia coli that has been shown to be important in genome maintenance. They function through catalyzing the reaction ATP + H2O → ADP + P and thus driving the unwinding of paired DNA and translocating in the 3' to 5' direction. These enzymes can also drive the reaction NTP + H2O → NDP + P to drive the unwinding of either DNA or RNA.
Spermatocytes are a type of male gametocyte in animals. They derive from immature germ cells called spermatogonia. They are found in the testis, in a structure known as the seminiferous tubules. There are two types of spermatocytes, primary and secondary spermatocytes. Primary and secondary spermatocytes are formed through the process of spermatocytogenesis.
Gene conversion is the process by which one DNA sequence replaces a homologous sequence such that the sequences become identical after the conversion event. Gene conversion can be either allelic, meaning that one allele of the same gene replaces another allele, or ectopic, meaning that one paralogous DNA sequence converts another.
A heteroduplex is a double-stranded (duplex) molecule of nucleic acid originated through the genetic recombination of single complementary strands derived from different sources, such as from different homologous chromosomes or even from different organisms.
Homologous recombination is a type of genetic recombination in which genetic information is exchanged between two similar or identical molecules of double-stranded or single-stranded nucleic acids.
Synapsis or Syzygy is the pairing of two chromosomes that occurs during meiosis. It allows matching-up of homologous pairs prior to their segregation, and possible chromosomal crossover between them. Synapsis takes place during prophase I of meiosis. When homologous chromosomes synapse, their ends are first attached to the nuclear envelope. These end-membrane complexes then migrate, assisted by the extranuclear cytoskeleton, until matching ends have been paired. Then the intervening regions of the chromosome are brought together, and may be connected by a protein-DNA complex called the synaptonemal complex. During synapsis, autosomes are held together by the synaptonemal complex along their whole length, whereas for sex chromosomes, this only takes place at one end of each chromosome.
Recombination hotspots are regions in a genome that exhibit elevated rates of recombination relative to a neutral expectation. The recombination rate within hotspots can be hundreds of times that of the surrounding region. Recombination hotspots result from higher DNA break formation in these regions, and apply to both mitotic and meiotic cells. This appellation can refer to recombination events resulting from the uneven distribution of programmed meiotic double-strand breaks.
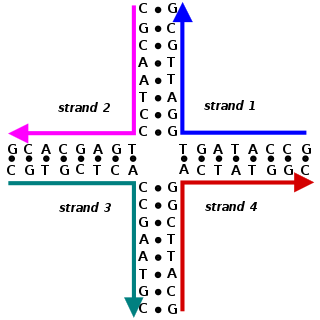
A Holliday junction is a branched nucleic acid structure that contains four double-stranded arms joined. These arms may adopt one of several conformations depending on buffer salt concentrations and the sequence of nucleobases closest to the junction. The structure is named after Robin Holliday, the molecular biologist who proposed its existence in 1964.
Mitotic recombination is a type of genetic recombination that may occur in somatic cells during their preparation for mitosis in both sexual and asexual organisms. In asexual organisms, the study of mitotic recombination is one way to understand genetic linkage because it is the only source of recombination within an individual. Additionally, mitotic recombination can result in the expression of recessive alleles in an otherwise heterozygous individual. This expression has important implications for the study of tumorigenesis and lethal recessive alleles. Mitotic homologous recombination occurs mainly between sister chromatids subsequent to replication. Inter-sister homologous recombination is ordinarily genetically silent. During mitosis the incidence of recombination between non-sister homologous chromatids is only about 1% of that between sister chromatids.
Sister chromatid exchange (SCE) is the exchange of genetic material between two identical sister chromatids.
A bivalent is one pair of chromosomes in a tetrad. A tetrad is the association of a pair of homologous chromosomes physically held together by at least one DNA crossover. This physical attachment allows for alignment and segregation of the homologous chromosomes in the first meiotic division. In most organisms, each replicated chromosome elicits formation of DNA double-strand breaks during the leptotene phase. These breaks are repaired by homologous recombination, that uses the homologous chromosome as a template for repair. The search for the homologous target, helped by numerous proteins collectively referred as the synaptonemal complex, cause the two homologs to pair, between the leptotene and the pachytene phases of meiosis I.
Chromosome segregation is the process in eukaryotes by which two sister chromatids formed as a consequence of DNA replication, or paired homologous chromosomes, separate from each other and migrate to opposite poles of the nucleus. This segregation process occurs during both mitosis and meiosis. Chromosome segregation also occurs in prokaryotes. However, in contrast to eukaryotic chromosome segregation, replication and segregation are not temporally separated. Instead segregation occurs progressively following replication.
The origin and function of meiosis are currently not well understood scientifically, and would provide fundamental insight into the evolution of sexual reproduction in eukaryotes. There is no current consensus among biologists on the questions of how sex in eukaryotes arose in evolution, what basic function sexual reproduction serves, and why it is maintained, given the basic two-fold cost of sex. It is clear that it evolved over 1.2 billion years ago, and that almost all species which are descendants of the original sexually reproducing species are still sexual reproducers, including plants, fungi, and animals.
Synthesis-dependent strand annealing (SDSA) is a major mechanism of homology-directed repair of DNA double-strand breaks (DSBs). Although many of the features of SDSA were first suggested in 1976, the double-Holliday junction model proposed in 1983 was favored by many researchers. In 1994, studies of double-strand gap repair in Drosophila were found to be incompatible with the double-Holliday junction model, leading researchers to propose a model they called synthesis-dependent strand annealing. Subsequent studies of meiotic recombination in S. cerevisiae found that non-crossover products appear earlier than double-Holliday junctions or crossover products, challenging the previous notion that both crossover and non-crossover products are produced by double-Holliday junctions and leading the authors to propose that non-crossover products are generated through SDSA.
Crossover interference is the term used to refer to the non-random placement of crossovers with respect to each other during meiosis. The term is attributed to Hermann Joseph Muller, who observed that one crossover "interferes with the coincident occurrence of another crossing over in the same pair of chromosomes, and I have accordingly termed this phenomenon ‘interference’."
↑ Bajrović K, Jevrić-Čaušević A, Hadžiselimović R, eds. (2005). Uvod u genetičko inženjerstvo i biotehnologiju. Institut za genetičko inženjerstvo i biotehnologiju (INGEB) Sarajevo. ISBN978-9958-9344-1-4.
↑ Alberts B (2002). Molecular Biology of the Cell, Fourth Edition. New York: Garland Science. ISBN978-0-8153-3218-3.
↑ "Access Excellence". Crossing-over: Genetic Recombination. The National Health Museum Resource Center. Retrieved February 23, 2011.
↑ King RC, Stransfield WD (1998). Dictionary of Genetics. New York, Oxford: Oxford University Press. ISBN0-19-509442-5.
↑ Stacey KA (1994). "Recombination". In Kendrew J, Lawrence E (eds.). The Encyclopedia of Molecular Biology. Oxford: Blackwell Science. pp.945–950.
↑ Boyd JB (1978). "DNA repair in Drosophila". In Hanawalt PC, Friedberg EC, Fox CF (eds.). DNA Repair Mechanisms. New York: Academic Press. pp.449–452.
↑ Galetzka D, Weis E, Kohlschmidt N, Bitz O, Stein R, Haaf T (April 2007). "Expression of somatic DNA repair genes in human testes". Journal of Cellular Biochemistry. 100 (5): 1232–9. doi:10.1002/jcb.21113. PMID17177185. S2CID23743474.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.