Related Research Articles

The entorhinal cortex (EC) is an area of the brain's allocortex, located in the medial temporal lobe, whose functions include being a widespread network hub for memory, navigation, and the perception of time. The EC is the main interface between the hippocampus and neocortex. The EC-hippocampus system plays an important role in declarative (autobiographical/episodic/semantic) memories and in particular spatial memories including memory formation, memory consolidation, and memory optimization in sleep. The EC is also responsible for the pre-processing (familiarity) of the input signals in the reflex nictitating membrane response of classical trace conditioning; the association of impulses from the eye and the ear occurs in the entorhinal cortex.
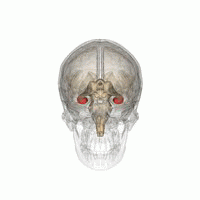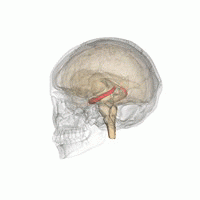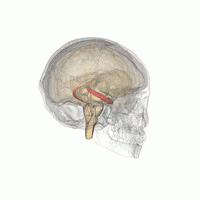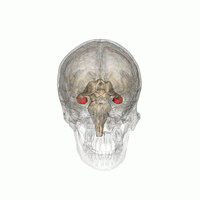
The hippocampus is a major component of the brain of humans and other vertebrates. Humans and other mammals have two hippocampi, one in each side of the brain. The hippocampus is part of the limbic system, and plays important roles in the consolidation of information from short-term memory to long-term memory, and in spatial memory that enables navigation. The hippocampus is located in the allocortex, with neural projections into the neocortex, in humans as well as other primates. The hippocampus, as the medial pallium, is a structure found in all vertebrates. In humans, it contains two main interlocking parts: the hippocampus proper, and the dentate gyrus.

A place cell is a kind of pyramidal neuron in the hippocampus that becomes active when an animal enters a particular place in its environment, which is known as the place field. Place cells are thought to act collectively as a cognitive representation of a specific location in space, known as a cognitive map. Place cells work with other types of neurons in the hippocampus and surrounding regions to perform this kind of spatial processing. They have been found in a variety of animals, including rodents, bats, monkeys and humans.
Explicit memory is one of the two main types of long-term human memory, the other of which is implicit memory. Explicit memory is the conscious, intentional recollection of factual information, previous experiences, and concepts. This type of memory is dependent upon three processes: acquisition, consolidation, and retrieval.
Schaffer collaterals are axon collaterals given off by CA3 pyramidal cells in the hippocampus. These collaterals project to area CA1 of the hippocampus and are an integral part of memory formation and the emotional network of the Papez circuit, and of the hippocampal trisynaptic loop. It is one of the most studied synapses in the world and named after the Hungarian anatomist-neurologist Károly Schaffer.
An apical dendrite is a dendrite that emerges from the apex of a pyramidal cell. Apical dendrites are one of two primary categories of dendrites, and they distinguish the pyramidal cells from spiny stellate cells in the cortices. Pyramidal cells are found in the prefrontal cortex, the hippocampus, the entorhinal cortex, the olfactory cortex, and other areas. Dendrite arbors formed by apical dendrites are the means by which synaptic inputs into a cell are integrated. The apical dendrites in these regions contribute significantly to memory, learning, and sensory associations by modulating the excitatory and inhibitory signals received by the pyramidal cells.
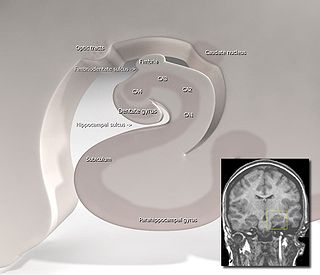
The subiculum is the most inferior component of the hippocampal formation. It lies between the entorhinal cortex and the CA1 subfield of the hippocampus proper.

Hippocampal sclerosis (HS) or mesial temporal sclerosis (MTS) is a neuropathological condition with severe neuronal cell loss and gliosis in the hippocampus. Neuroimaging tests such as magnetic resonance imaging (MRI) and positron emission tomography (PET) may identify individuals with hippocampal sclerosis. Hippocampal sclerosis occurs in 3 distinct settings: mesial temporal lobe epilepsy, adult neurodegenerative disease and acute brain injury.
Theta waves generate the theta rhythm, a neural oscillation in the brain that underlies various aspects of cognition and behavior, including learning, memory, and spatial navigation in many animals. It can be recorded using various electrophysiological methods, such as electroencephalogram (EEG), recorded either from inside the brain or from electrodes attached to the scalp.

In the field of neurology, temporal lobe epilepsy is an enduring brain disorder that causes unprovoked seizures from the temporal lobe. Temporal lobe epilepsy is the most common type of focal onset epilepsy among adults. Seizure symptoms and behavior distinguish seizures arising from the medial temporal lobe from seizures arising from the lateral (neocortical) temporal lobe. Memory and psychiatric comorbidities may occur. Diagnosis relies on electroencephalographic (EEG) and neuroimaging studies. Anticonvulsant medications, epilepsy surgery and dietary treatments may improve seizure control.
In the brain, the perforant path or perforant pathway provides a connectional route from the entorhinal cortex to all fields of the hippocampal formation, including the dentate gyrus, all CA fields, and the subiculum.

Michael Hasselmo is an American neuroscientist and professor in the Department of Psychological and Brain Sciences at Boston University. He is the director of the Center for Systems Neuroscience and is editor-in-chief of Hippocampus (journal). Hasselmo studies oscillatory dynamics and neuromodulatory regulation in cortical mechanisms for memory guided behavior and spatial navigation using a combination of neurophysiological and behavioral experiments in conjunction with computational modeling. In addition to his peer-reviewed publications, Hasselmo wrote the book How We Remember: Brain Mechanisms of Episodic Memory.
The trisynaptic circuit or trisynaptic loop is a relay of synaptic transmission in the hippocampus. The circuit was initially described by the neuroanatomist Santiago Ramon y Cajal, in the early twentieth century, using the Golgi staining method. After the discovery of the trisynaptic circuit, a series of research has been conducted to determine the mechanisms driving this circuit. Today, research is focused on how this loop interacts with other parts of the brain, and how it influences human physiology and behaviour. For example, it has been shown that disruptions within the trisynaptic circuit lead to behavioural changes in rodent and feline models.

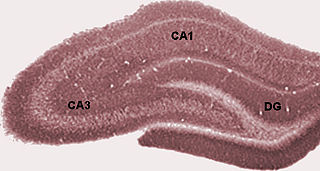
Hippocampus anatomy describes the physical aspects and properties of the hippocampus, a neural structure in the medial temporal lobe of the brain. It has a distinctive, curved shape that has been likened to the sea-horse monster of Greek mythology and the ram's horns of Amun in Egyptian mythology. This general layout holds across the full range of mammalian species, from hedgehog to human, although the details vary. For example, in the rat, the two hippocampi look similar to a pair of bananas, joined at the stems. In primate brains, including humans, the portion of the hippocampus near the base of the temporal lobe is much broader than the part at the top. Due to the three-dimensional curvature of this structure, two-dimensional sections such as shown are commonly seen. Neuroimaging pictures can show a number of different shapes, depending on the angle and location of the cut.
Memory consolidation is a category of processes that stabilize a memory trace after its initial acquisition. A memory trace is a change in the nervous system caused by memorizing something. Consolidation is distinguished into two specific processes. The first, synaptic consolidation, which is thought to correspond to late-phase long-term potentiation, occurs on a small scale in the synaptic connections and neural circuits within the first few hours after learning. The second process is systems consolidation, occurring on a much larger scale in the brain, rendering hippocampus-dependent memories independent of the hippocampus over a period of weeks to years. Recently, a third process has become the focus of research, reconsolidation, in which previously consolidated memories can be made labile again through reactivation of the memory trace.

The relationship between sleep and memory has been studied since at least the early 19th century. Memory, the cognitive process of storing and retrieving past experiences, learning and recognition, is a product of brain plasticity, the structural changes within synapses that create associations between stimuli. Stimuli are encoded within milliseconds; however, the long-term maintenance of memories can take additional minutes, days, or even years to fully consolidate and become a stable memory that is accessible. Therefore, the formation of a specific memory occurs rapidly, but the evolution of a memory is often an ongoing process.
Sharp waves and ripples (SWRs) are oscillatory patterns produced by extremely synchronised activity of neurons in the mammalian hippocampus and neighbouring regions which occur spontaneously in idle waking states or during NREM sleep. They can be observed with a variety of imaging methods, such as EEG. They are composed of large amplitude sharp waves in local field potential and produced by tens of thousands of neurons firing together within 30–100 ms window. They are some of the most synchronous oscillations patterns in the brain, making them susceptible to pathological patterns such as epilepsy.They have been extensively characterised and described by György Buzsáki and have been shown to be involved in memory consolidation in NREM sleep and the replay of memories acquired during wakefulness.
Chantal Stern is a neuroscientist who uses techniques including functional magnetic resonance imaging (fMRI) to study the brain mechanisms of memory function. She is the Director of the Brain, Behavior and Cognition program and a professor of Psychological and Brain Sciences at Boston University.After completing a degree at McGill University, she performed her doctoral research at Oxford University with Richard Passingham.
The hippocampus proper refers to the actual structure of the hippocampus which is made up of three regions or subfields. The subfields CA1, CA2, and CA3 use the initials of cornu Ammonis, an earlier name of the hippocampus.
Early long-term potentiation (E-LTP) is the first phase of long-term potentiation (LTP), a well-studied form of synaptic plasticity, and consists of an increase in synaptic strength. LTP could be produced by repetitive stimulation of the presynaptic terminals, and it is believed to play a role in memory function in the hippocampus, amygdala and other cortical brain structures in mammals.
References
- 1 2 3 4 5 6 Gazzaniga, Michael S., Richard B. Ivry, and G. R. Mangun. "Chapter 9: Memory." Cognitive Neuroscience: The Biology of the Mind. 4th ed. New York: W. W. Norton, 2014. 378-423. Print.
- 1 2 3 4 5 Lepage, M.; Habib, R.; Tulving, E. (1998). "Lepage, M., Habib, R. & Tulving, E. Hippocampal PET Activations of memory encoding and retrieval: the HIPER model" Hippocampus 8, 313-322". Hippocampus. 8 (4): 313–22. doi:10.1002/(SICI)1098-1063(1998)8:4<313::AID-HIPO1>3.0.CO;2-I. PMID 9744418. S2CID 14507536.
- 1 2 3 4 5 Hasselmo, ME; Bodelon, C; Wyble, BP (2002). "A proposed function for hippocampal theta rhythm: separate phases of encoding and retrieval enhance reversal of prior learning". Neural Computation. 14 (4): 793–817. doi:10.1162/089976602317318965. PMID 11936962. S2CID 9128504.
- 1 2 3 4 Kunec, S; Hasselmo, ME; Kopell, N (2005). "Encoding and Retrieval in the CA3 Region of the Hippocampus: A Model of Theta-Phase Separation". Journal of Neurophysiology. 94 (1): 70–82. CiteSeerX 10.1.1.333.5452 . doi:10.1152/jn.00731.2004. PMID 15728768.
- 1 2 3 Morris, R. G. M.; Inglis, J.; Ainge, J. A.; Olverman, H. J.; Tulloch, J.; Dudai, Y.; Kelly, P. A. T. (2006). "Memory reconsolidation: Sensitivity of spatial memory to inhibition of protein synthesis in dorsal hippocampus during encoding and retrieval". Neuron. 50 (3): 479–489. doi: 10.1016/j.neuron.2006.04.012 . PMID 16675401.
- 1 2 3 Nader, Karim; Schafe, Glenn E.; Le Doux, Joseph E. (2000). "Fear Memories Require Protein Synthesis In The Amygdala For Reconsolidation After Retrieval". Nature. 406 (6797): 722–726. Bibcode:2000Natur.406..722N. doi:10.1038/35021052. PMID 10963596. S2CID 4420637.
- 1 2 3 Bremner, J.; Vythilingam, M.; Vermetten, E.; Southwick, S.; MaGlashan, T.; Nazeer, A.; Khan, S.; Vaccarino, V.; Soufer, R.; Garg, P.; Chin, K.; Staib, L.; Duncan, J.; Charney, D. (2003). "MRI and PET study of deficits in hippocampal structure and function in women with childhood sexual abuse and posttraumatic stress disorder". The American Journal of Psychiatry. 160 (5): 924–932. doi:10.1176/appi.ajp.160.5.924. PMID 12727697.
- 1 2 Tan, F. (2014) Epilepsy and memory. BC Epilepsy Society, 1-12
- 1 2 3 Johns, P., Thom, M. (2008) Epilepsy and hippocampal sclerosis: cause or effect? Neuropathology Article, 8, 16-18
- ↑ Du, A.; Schuff, N.; Amend, D.; Laakso, M.; Hsu, Y.; Jagust, W.; Yaffe, K.; Kramer, J.; Reed, B.; Norman, D.; Chui, H.; Weiner, M. (2001). "Magnetic resonance imaging of the entorhinal cortex and hippocampus in mild cognitive impairment and alzheimer's disease". Journal of Neurology, Neurosurgery, and Psychiatry. 71 (4): 441–447. doi:10.1136/jnnp.71.4.441. PMC 1763497 . PMID 11561025.
- 1 2 3 4 Zeineh, M (2003). "Dynamics of the Hippocampus During Encoding and Retrieval of Face-Name Pairs". Science. 299 (5606): 577–580. Bibcode:2003Sci...299..577Z. doi:10.1126/science.1077775. PMID 12543980. S2CID 2361898.