Related Research Articles

The visual cortex of the brain is the area of the cerebral cortex that processes visual information. It is located in the occipital lobe. Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex. The area of the visual cortex that receives the sensory input from the lateral geniculate nucleus is the primary visual cortex, also known as visual area 1 (V1), Brodmann area 17, or the striate cortex. The extrastriate areas consist of visual areas 2, 3, 4, and 5.
Blindsight is the ability of people who are cortically blind due to lesions in their striate cortex, also known as the primary visual cortex or V1, to respond to visual stimuli that they do not consciously see.

The lateral geniculate nucleus is a relay center in the thalamus for the visual pathway. It receives a major sensory input from the retina. The LGN is the main central connection for the optic nerve to the occipital lobe, particularly the primary visual cortex. In humans, each LGN has six layers of neurons alternating with optic fibers.

The parietal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The parietal lobe is positioned above the temporal lobe and behind the frontal lobe and central sulcus.
The pulvinar nuclei or nuclei of the pulvinar are the nuclei located in the thalamus. As a group they make up the collection called the pulvinar of the thalamus, usually just called the pulvinar.

The superior colliculus is a structure lying on the roof of the mammalian midbrain. In non-mammalian vertebrates, the homologous structure is known as the optic tectum, or optic lobe. The adjective form tectal is commonly used for both structures.
The pretectal area, or pretectum, is a midbrain structure composed of seven nuclei and comprises part of the subcortical visual system. Through reciprocal bilateral projections from the retina, it is involved primarily in mediating behavioral responses to acute changes in ambient light such as the pupillary light reflex, the optokinetic reflex, and temporary changes to the circadian rhythm. In addition to the pretectum's role in the visual system, the anterior pretectal nucleus has been found to mediate somatosensory and nociceptive information.
Multisensory integration, also known as multimodal integration, is the study of how information from the different sensory modalities may be integrated by the nervous system. A coherent representation of objects combining modalities enables animals to have meaningful perceptual experiences. Indeed, multisensory integration is central to adaptive behavior because it allows animals to perceive a world of coherent perceptual entities. Multisensory integration also deals with how different sensory modalities interact with one another and alter each other's processing.
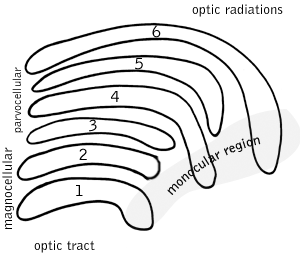
A koniocellular cell is a neuron with a small cell body that is located in the koniocellular layer of the lateral geniculate nucleus (LGN) in primates, including humans.
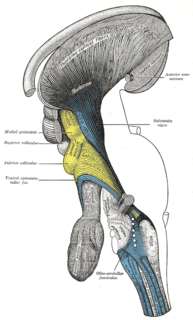
Thalamocortical radiations are the fibers between the thalamus and the cerebral cortex.
The zona incerta (ZI) is a horizontally elongated region of gray matter in the subthalamus below the thalamus. Its connections project extensively over the brain from the cerebral cortex down into the spinal cord.

The frontal eye fields (FEF) are a region located in the frontal cortex, more specifically in Brodmann area 8 or BA8, of the primate brain. In humans, it can be more accurately said to lie in a region around the intersection of the middle frontal gyrus with the precentral gyrus, consisting of a frontal and parietal portion. The FEF is responsible for saccadic eye movements for the purpose of visual field perception and awareness, as well as for voluntary eye movement. The FEF communicates with extraocular muscles indirectly via the paramedian pontine reticular formation. Destruction of the FEF causes deviation of the eyes to the ipsilateral side.

Supplementary eye field (SEF) is the name for the anatomical area of the dorsal medial frontal lobe of the primate cerebral cortex that is indirectly involved in the control of saccadic eye movements. Evidence for a supplementary eye field was first shown by Schlag, and Schlag-Rey. Current research strives to explore the SEF's contribution to visual search and its role in visual salience. The SEF constitutes together with the frontal eye fields (FEF), the intraparietal sulcus (IPS), and the superior colliculus (SC) one of the most important brain areas involved in the generation and control of eye movements, particularly in the direction contralateral to their location. Its precise function is not yet fully known. Neural recordings in the SEF show signals related to both vision and saccades somewhat like the frontal eye fields and superior colliculus, but currently most investigators think that the SEF has a special role in high level aspects of saccade control, like complex spatial transformations, learned transformations, and executive cognitive functions.

The medial dorsal nucleus is a large nucleus in the thalamus.
The posterior parietal cortex plays an important role in planned movements, spatial reasoning, and attention.
Inferior pulvinar nucleus is one of four traditionally anatomically distinguished nuclei of the pulvinar of the thalamus. The other three nuclei of the pulvinar are called lateral, anterior and medial pulvinar nuclei.
Lateral pulvinar nucleus is one of four traditionally anatomically distinguished nuclei of the pulvinar of the thalamus. The other three nuclei of the pulvinar are called anterior, inferior and medial pulvinar nuclei.
Medial pulvinar nucleus is one of four traditionally anatomically distinguished nuclei of the pulvinar of the thalamus. The other three nuclei of the pulvinar are called lateral, inferior and anterior pulvinar nuclei.

Peter H. Schiller is a professor emeritus of Neuroscience in the Department of Brain and Cognitive Sciences at the Massachusetts Institute of Technology (MIT). He is well known for his work on the behavioral, neurophysiological and pharmacological studies of the primate visual and oculomotor systems.
Michael E. Goldberg, also known as Mickey Goldberg, is an American neuroscientist and David Mahoney Professor at Columbia University. He is known for his work on the mechanisms of the mammalian eye in relation to brain activity. He served as president of the Society for Neuroscience from 2009 to 2010.
References
- 1 2 3 Whishaw, I. Q., & Kolb, B. (2015). Fundamentals of human neuropsychology. New York, NY: Worth Custom Publishing.
- 1 2 3 4 Banich, M. T., & Compton, R. J. (2018). Cognitive neuroscience. Cambridge, United Kingdom: Cambridge University Press.
- ↑ Nagel, S. M. (n.d.). Superior Colliculus. Retrieved November 20, 2019, from https://psych.athabascau.ca/html/Psych402/Biotutorials/24/colliculus.shtml
- ↑ Visual cortex. (n.d.). In Wikipedia. Retrieved November 20, 2019, from https://en.wikipedia.org/wiki/Visual_cortex.
- 1 2 Pulvinar nuclei. (n.d.). In Wikipedia. Retrieved November 20, 2019, from https://en.wikipedia.org/wiki/Pulvinar_nuclei.
- ↑ Swearer J. (2011) Optic Ataxia. In: Kreutzer J.S., DeLuca J., Caplan B. (eds) Encyclopedia of Clinical Neuropsychology. Springer, New York, NY
- ↑ Leh, S. E., Johansen-Berg, H., & Ptito, A. (2006). Unconscious vision: new insights into the neuronal correlate of blindsight using diffusion tractography. Brain, 129(7), 1822–1832. doi: 10.1093/brain/awl111
- ↑ Mcpeek, R. M., & Keller, E. L. (2010). Deficits in saccade target selection after temporary inactivation of superior colliculus. Journal of Vision, 2(7), 573–573. doi: 10.1167/2.7.573
- ↑ Krauzlis, R. J., Lovejoy, L. P., & Zénon, A. (2013). Superior Colliculus and Visual Spatial Attention. Annual Review of Neuroscience, 36(1), 165–182. doi: 10.1146/annurev-neuro-062012-170249
- ↑ Song, J.-H., & Mcpeek, R. M. (2015). Neural correlates of target selection for reaching movements in superior colliculus. Journal of Neurophysiology, 113(5), 1414–1422. doi: 10.1152/jn.00417.2014
- ↑ Ghose, D., Maier, A., Nidiffer, A., & Wallace, M. T. (2014). Multisensory Response Modulation in the Superficial Layers of the Superior Colliculus. Journal of Neuroscience, 34(12), 4332–4344. doi: 10.1523/jneurosci.3004-13.2014
- ↑ Petersen, S. E., Robinson, D. L., & Morris, J. (1987). Contributions of the pulvinar to visual spatial attention. Neuropsychologia, 25(1), 97–105. doi: 10.1016/0028-3932(87)90046-7