
Euryoryzomys emmonsae, also known as Emmons' rice rat or Emmons' oryzomys, is a rodent from the Amazon rainforest of Brazil in the genus Euryoryzomys of the family Cricetidae. Initially misidentified as E. macconnelli or E. nitidus, it was formally described in 1998. A rainforest species, it may be scansorial, climbing but also spending time on the ground. It lives only in a limited area south of the Amazon River in the state of Pará, a distribution that is apparently unique among the muroid rodents of the region.

Lundomys molitor, also known as Lund's amphibious rat or the greater marsh rat, is a semiaquatic rat species from southeastern South America.

Pseudoryzomys simplex, also known as the Brazilian false rice rat or false oryzomys, is a species of rodent in the family Cricetidae from south-central South America. It is found in lowland palm savanna and thorn scrub habitats. It is a medium-sized species, weighing about 50 grams (1.8 oz), with gray–brown fur, long and narrow hindfeet, and a tail that is about as long as the head and body. The IUCN has assessed its conservation status as being of least concern, although almost nothing is known about its diet or reproduction.

Transandinomys bolivaris, also known as the long-whiskered rice rat, is a rodent in the family Cricetidae. It is found in humid forest from northeastern Honduras to western Ecuador, up to 1,800 m (5,900 ft) above sea level. Since it was first described in 1901 from Ecuador, six scientific names have been introduced for it, but their common identity was not documented until 1998 and the species has long been known under the name Oryzomys bombycinus, described from Panama in 1912. The name Oryzomys bolivaris was used before it was moved to the new genus Transandinomys with Transandinomys talamancae in 2006.

Oryzomys gorgasi, also known as Gorgas's oryzomys or Gorgas's rice rat, is a rodent in the genus Oryzomys of family Cricetidae. First recorded in 1967, it is known from only a few localities, including a freshwater swamp in the lowlands of northwestern Colombia and a mangrove islet in northwestern Venezuela. It reportedly formerly occurred on the island of Curaçao off northwestern Venezuela; this extinct population has been described as a separate species, Oryzomys curasoae, but does not differ morphologically from mainland populations.
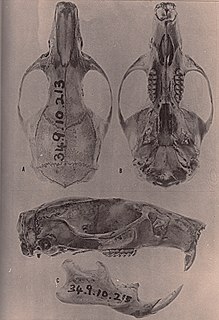
Mindomys hammondi, also known as Hammond's rice rat or Hammond's oryzomys, is an endangered species of rodent in the tribe Oryzomyini of family Cricetidae. Formerly considered to be related with Nectomys, Sigmodontomys, Megalomys, or Oryzomys, it is now placed in then genus Mindomys, but its relationships remain obscure; some evidence supports a placement near Oecomys or as a basal member of Oryzomyini.

Eremoryzomys polius, also known as the gray rice rat or the Marañon oryzomys, is a rodent species in the tribe Oryzomyini of the family Cricetidae. Discovered in 1912 and first described in 1913 by Wilfred Osgood, it was originally placed in Oryzomys and named Oryzomys polius. In 2006, a cladistic analysis found that it was not closely related to Oryzomys in the strict sense or to any other oryzomyine then known, so that it is now placed in its own genus, Eremoryzomys. The Brazilian genus Drymoreomys, named in 2011, is probably the closest relative of Eremoryzomys. Eremoryzomys has a limited distribution in the dry upper valley of the Marañón River in central Peru, but may yet contain more than one species.

Oryzomyini is a tribe of rodents in the subfamily Sigmodontinae of the family Cricetidae. It includes about 120 species in about thirty genera, distributed from the eastern United States to the southernmost parts of South America, including many offshore islands. It is part of the clade Oryzomyalia, which includes most of the South American Sigmodontinae.

Noronhomys vespuccii, also known as Vespucci's rodent, is an extinct rat species from the islands of Fernando de Noronha off northeastern Brazil. Italian explorer Amerigo Vespucci may have seen it on a visit to Fernando de Noronha in 1503, but it subsequently became extinct, perhaps because of the exotic rats and mice introduced by the first explorers of the island. Numerous but fragmentary fossil remains of the animal, of uncertain but probably Holocene age, were discovered in 1973 and described in 1999.
Reigomys primigenus is an extinct oryzomyine rodent known from Pleistocene deposits in Tarija Department, southeastern Bolivia. It is known from a number of isolated jaws and molars which show that its molars were almost identical to those of the living Lundomys. On the other hand, the animal possesses a number of derived traits of the palate which document a closer relationship to living Holochilus, the genus of South American marsh rats, and for this reason it was placed in the genus Holochilus when it was first described in 1996. The subsequent discoveries of Noronhomys and Carletonomys, which may be more closely related to extant Holochilus than H. primigenus is, have cast its placement in Holochilus into doubt, and it was ultimately made the type species of a separate genus, Reigomys.

Transandinomys is a genus of rodents in the tribe Oryzomyini of family Cricetidae. It includes two species—T. bolivaris and T. talamancae—found in forests from Honduras in Central America south and east to southwestern Ecuador and northwestern Venezuela in northern South America. Until 2006, its members were included in the genus Oryzomys, but phylogenetic analysis showed that they are not closely related to the type species of that genus, and they have therefore been placed in a new genus. They may be most closely related to genera like Hylaeamys and Euryoryzomys, which contain very similar species. Both species of Transandinomys have had eventful taxonomic histories.

Nephelomys is a genus of South American oryzomyine rodents found in the Andes from Bolivia to Venezuela, with a westward extension into the mountains of Costa Rica. Its generic name is derived from the Ancient Greek word nephelê "mist", referring to the cloud forest habitat of the members of the genus.
In anatomy, posterolateral palatal pits are gaps at the sides of the back of the bony palate, near the last molars. Posterolateral palatal pits are present, in various degrees of development, in several members of the rodent family Cricetidae. Many members of the family lack them or have only simple pits, but Arvicolinae and Oryzomyini have more highly developed posterolateral palatal pits. Posterolateral palatal pits are also present in some other rodents, including Glis, Jaculus, Hystrix, Abrocoma, Ctenomys, Chinchilla, and Lagidium.

In rodent anatomy, the zygomatic plate is a bony plate derived from the flattened front part of the zygomatic arch (cheekbone). At the back, it connects to the front (maxillary) root of the zygomatic arch, and at the top it is connected to the rest of the skull via the antorbital bridge. It is part of the maxillary bone, or upper jaw, which also contains the upper cheekteeth. Primitively, rodents have a nearly horizontal zygomatic plate. In association with specializations in zygomasseteric system, several distinct morphologies have developed across the order.
In rodents, sphenopalatine vacuities are perforations of the roof of the mesopterygoid fossa, the open space behind the palate, in between the parapterygoid fossae. They may perforate the presphenoid or basisphenoid bone. Their development and form are variable between and within species, and features of the sphenopalatine vacuities have been used as characters in cladistic analyses.
In rodents, incisor procumbency refers to the orientation of the upper incisor, defined by the position of the cutting edge of the incisor relative to the vertical plane of the incisors. Proodont incisors have the cutting edge in front of the vertical plane, orthodont teeth have it perpendicular to the plane, opisthodont incisors have it behind the plane, and hyper-opisthodont teeth have the cutting edge even behind the back of the alveolus of the incisor.
In some rodents, squamae are small tubercles resembling scales on the sole of the hindfeet. Among oryzomyine rodents, their development is variable; most have well-developed squamae, but in others they are indistinct or entirely absent. Delomys sublineatus and Peromyscus maniculatus also have squamae.

Agathaeromys is an extinct genus of oryzomyine rodents from the Pleistocene of Bonaire, Netherlands Antilles. Two species are known, which differ in size and some details of tooth morphology. The larger A. donovani, the type species, is known from hundreds of teeth that are probably 900,000 to 540,000 years old, found in four localities. A. praeuniversitatis, the smaller species, is known from 35 teeth found in a single fossil site, which is probably 540,000 to 230,000 years old.
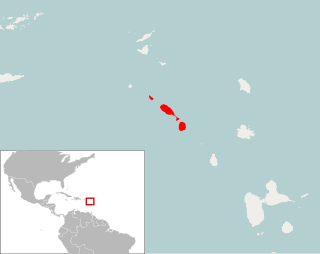
Pennatomys nivalis is an extinct oryzomyine rodent from the islands of Sint Eustatius, Saint Kitts, and Nevis in the Lesser Antilles. The only species in the genus Pennatomys, it is known from skeletal remains found in Amerindian archeological sites on all three islands, with dates ranging from 790–520 BCE to 900–1200 CE. No live specimens are known, but there are several historical records of rodents from Saint Kitts and Nevis that could conceivably refer to Pennatomys. The animal apparently belongs to a group within the tribe Oryzomyini that includes many other island-dwelling species.

Drymoreomys is a rodent genus in the tribe Oryzomyini that lives in the Atlantic Forest of Brazil. The single species, D. albimaculatus, is known only from the states of São Paulo and Santa Catarina and was not named until 2011. It lives in the humid forest on the eastern slopes of the Serra do Mar and perhaps reproduces year-round. Although its range is relatively large and includes some protected areas, it is patchy and threatened, and the discoverers recommend that the animal be considered "Near Threatened" on the IUCN Red List. Within Oryzomyini, Drymoreomys appears to be most closely related to Eremoryzomys from the Andes of Peru, a biogeographically unusual relationship, in that the two populations are widely separated and each is adapted to an arid or a moist environment.
