Related Research Articles

A cactus is a member of the plant family Cactaceae, a family comprising about 127 genera with some 1750 known species of the order Caryophyllales. The word "cactus" derives, through Latin, from the Ancient Greek κάκτος, kaktos, a name originally used by Theophrastus for a spiny plant whose identity is now not certain. Cacti occur in a wide range of shapes and sizes. Most cacti live in habitats subject to at least some drought. Many live in extremely dry environments, even being found in the Atacama Desert, one of the driest places on earth. Cacti show many adaptations to conserve water. Almost all cacti are succulents, meaning they have thickened, fleshy parts adapted to store water. Unlike many other succulents, the stem is the only part of most cacti where this vital process takes place. Most species of cacti have lost true leaves, retaining only spines, which are highly modified leaves. As well as defending against herbivores, spines help prevent water loss by reducing air flow close to the cactus and providing some shade. In the absence of leaves, enlarged stems carry out photosynthesis. Cacti are native to the Americas, ranging from Patagonia in the south to parts of western Canada in the north—except for Rhipsalis baccifera, which also grows in Africa and Sri Lanka.

Plant cells are eukaryotic cells present in green plants, photosynthetic eukaryotes of the kingdom Plantae. Their distinctive features include primary cell walls containing cellulose, hemicelluloses and pectin, the presence of plastids with the capability to perform photosynthesis and store starch, a large vacuole that regulates turgor pressure, the absence of flagella or centrioles, except in the gametes, and a unique method of cell division involving the formation of a cell plate or phragmoplast that separates the new daughter cells.
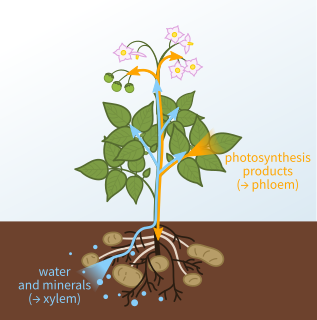
Xylem is one of the two types of transport tissue in vascular plants, the other being phloem. The basic function of xylem is to transport water from roots to stems and leaves, but it also transports nutrients. The word "xylem" is derived from the Greek word ξύλον (xylon), meaning "wood"; the best-known xylem tissue is wood, though it is found throughout a plant. The term was introduced by Carl Nägeli in 1858.

In vascular plants, the roots are the organs of a plant that are modified to provide anchorage for the plant and take in water and nutrients into the plant body, which allows plants to grow taller and faster. They most often lie below the surface of the soil, but roots can also be aerial or aerating, that is, growing up above the ground or especially above water.

Vascular plants, also known as Tracheophyta, form a large group of plants that are defined as land plants with lignified tissues for conducting water and minerals throughout the plant. They also have a specialized non-lignified tissue to conduct products of photosynthesis. Vascular plants include the clubmosses, horsetails, ferns, gymnosperms and angiosperms. Scientific names for the group include Tracheophyta, Tracheobionta and Equisetopsida sensu lato. Some early land plants had less developed vascular tissue; the term eutracheophyte has been used for all other vascular plants.

In biology, tissue is a cellular organizational level between cells and a complete organ. A tissue is an ensemble of similar cells and their extracellular matrix from the same origin that together carry out a specific function. Organs are then formed by the functional grouping together of multiple tissues.

Bark is the outermost layers of stems and roots of woody plants. Plants with bark include trees, woody vines, and shrubs. Bark refers to all the tissues outside the vascular cambium and is a nontechnical term. It overlays the wood and consists of the inner bark and the outer bark. The inner bark, which in older stems is living tissue, includes the innermost layer of the periderm. The outer bark on older stems includes the dead tissue on the surface of the stems, along with parts of the outermost periderm and all the tissues on the outer side of the periderm. The outer bark on trees which lies external to the living periderm is also called the rhytidome.

The vascular cambium is the main growth tissue in the stems and roots of many plants, specifically in dicots such as buttercups and oak trees, gymnosperms such as pine trees, as well as in certain vascular plants. It produces secondary xylem inwards, towards the pith, and secondary phloem outwards, towards the bark.
In a vascular plant, the stele is the central part of the root or stem containing the tissues derived from the procambium. These include vascular tissue, in some cases ground tissue (pith) and a pericycle, which, if present, defines the outermost boundary of the stele. Outside the stele lies the endodermis, which is the innermost cell layer of the cortex.
Lepidodendron is an extinct genus of primitive, vascular, tree-like plants, also known as scale trees, related to the quillworts and lycopsids. They were part of the coal forest flora. They sometimes reached heights of 50 metres, and the trunks were often over 1 m (3.3 ft) in diameter. They thrived during the Carboniferous Period before going extinct. Sometimes erroneously called "giant club mosses", the genus was actually more closely related to modern quillworts than to modern club mosses.

Equisetidae is one of the four subclasses of Polypodiopsida (ferns), a group of vascular plants with a fossil record going back to the Devonian. They are commonly known as horsetails. They typically grow in wet areas, with whorls of needle-like branches radiating at regular intervals from a single vertical stem.

The ground tissue of plants includes all tissues that are neither dermal nor vascular. It can be divided into three types based on the nature of the cell walls.
- Parenchyma cells have thin primary walls and usually remain alive after they become mature. Parenchyma forms the "filler" tissue in the soft parts of plants, and is usually present in cortex, pericycle, pith, and medullary rays in primary stem and root.
- Collenchyma cells have thin primary walls with some areas of secondary thickening. Collenchyma provides extra mechanical and structural support, particularly in regions of new growth.
- Sclerenchyma cells have thick lignified secondary walls and often die when mature. Sclerenchyma provides the main structural support to a plant.

Vascular tissue is a complex conducting tissue, formed of more than one cell type, found in vascular plants. The primary components of vascular tissue are the xylem and phloem. These two tissues transport fluid and nutrients internally. There are also two meristems associated with vascular tissue: the vascular cambium and the cork cambium. All the vascular tissues within a particular plant together constitute the vascular tissue system of that plant.

In botany, secondary growth is the growth that results from cell division in the cambia or lateral meristems and that causes the stems and roots to thicken, while primary growth is growth that occurs as a result of cell division at the tips of stems and roots, causing them to elongate, and gives rise to primary tissue. Secondary growth occurs in most seed plants, but monocots usually lack secondary growth. If they do have secondary growth, it differs from the typical pattern of other seed plants.

Opuntioideae is a subfamily of the cactus family, Cactaceae. It contains 15 genera divided into five tribes. The subfamily encompasses roughly 220-250 species, and is geographically distributed throughout the New World from Canada, to Argentina. Members of this subfamily have diverse habits, including small geophytes, hemispherical cushions, shrubs, trees, and columnar cacti consisting of indeterminate branches or determinate terete or spherical segments.
Important structures in plant development are buds, shoots, roots, leaves, and flowers; plants produce these tissues and structures throughout their life from meristems located at the tips of organs, or between mature tissues. Thus, a living plant always has embryonic tissues. By contrast, an animal embryo will very early produce all of the body parts that it will ever have in its life. When the animal is born, it has all its body parts and from that point will only grow larger and more mature. However, both plants and animals pass through a phylotypic stage that evolved independently and that causes a developmental constraint limiting morphological diversification.

Lepidodendrales were primitive, vascular, arborescent (tree-like) plants related to the lycopsids. Members of Lepidodendrales are the best understood of the fossil lycopsids due to the vast diversity of Lepidodendrales specimens and the diversity in which they were preserved; the extensive distribution of Lepidodendrales specimens as well as their well-preservedness lends paleobotanists exceptionally detailed knowledge of the coal-swamp giants’ reproductive biology, vegetative development, and role in their paleoecosystem. The defining characteristics of the Lepidodendrales are their secondary xylem, extensive periderm development, three-zoned cortex, rootlike appendages known as stigmarian rootlets arranged in a spiralling pattern, and megasporangium each containing a single functional megaspore that germinates inside the sporangium. Many of these different plant organs have been assigned both generic and specific names as relatively few have been found organically attached to each other. Some specimens have been discovered which indicate heights of 40 and even 50 meters and diameters of over 2 meters at the base. The massive trunks of some species branched profusely, producing large crowns of leafy twigs; though some leaves were up to 1 meter long, most were much shorter, and when leaves dropped from branches their conspicuous leaf bases remained on the surface of branches. Strobili could be found at the tips of distal branches or in an area at the top of the main trunk. The underground organs of Lepidodendrales typically consisted of dichotomizing axes bearing helically arranged, lateral appendages serving an equivalent function to roots. Sometimes called "giant club mosses", they are in fact more closely related to quillworts than to club mosses.

A stem is one of two main structural axes of a vascular plant, the other being the root. It supports leaves, flowers and fruits, transports water and dissolved substances between the roots and the shoots in the xylem and phloem, stores nutrients, and produces new living tissue.

A leaf is the principal lateral appendage of the vascular plant stem, usually borne above ground and specialized for photosynthesis. The leaves, stem, flower and fruit together form the shoot system. Leaves are collectively referred to as foliage, as in "autumn foliage". In most leaves, the primary photosynthetic tissue, the palisade mesophyll, is located on the upper side of the blade or lamina of the leaf but in some species, including the mature foliage of Eucalyptus, palisade mesophyll is present on both sides and the leaves are said to be isobilateral. Most leaves are flattened and have distinct upper (adaxial) and lower (abaxial) surfaces that differ in color, hairiness, the number of stomata, the amount and structure of epicuticular wax and other features. Leaves are mostly green in color due to the presence of a compound called chlorophyll that is essential for photosynthesis as it absorbs light energy from the sun. A leaf with lighter-colored or white patches or edges is called a variegated leaf.

In plant morphology, thorns, spines, and prickles, and in general spinose structures, are hard, rigid extensions or modifications of leaves, roots, stems or buds with sharp, stiff ends, and generally serve the same function: physically deterring animals from eating the plant material.
References
- ↑ The particularity of the Cactus Cortex, Cactus Art.
- ↑ Transverse section of cactus stem. Plant Anatomy, James D. Mauseth.
- ↑ Cortical Bundles in the Persistent, Photosynthetic Stems of Cacti. Annals of Botany, Volume 70, Issue 4, pg 317-324.
| | This Cactaceae article is a stub. You can help Wikipedia by expanding it. |