
An RNA virus is a virus which has RNA as its genetic material. The nucleic acid is usually single-stranded RNA (ssRNA) but it may be double-stranded RNA (dsRNA). Notable human diseases caused by RNA viruses include the common cold, influenza, SARS, MERS, COVID-19, Dengue Virus, hepatitis C, hepatitis E, West Nile fever, Ebola virus disease, rabies, polio, mumps, and measles.

Defective interfering particles (DIPs), also known as defective interfering viruses, are spontaneously generated virus mutants in which a critical portion of the particle's genome has been lost due to defective replication or non-homologous recombination. The mechanism of their formation is presumed to be as a result of template-switching during replication of the viral genome, although non-replicative mechanisms involving direct ligation of genomic RNA fragments have also been proposed. DIPs are derived from and associated with their parent virus, and particles are classed as DIPs if they are rendered non-infectious due to at least one essential gene of the virus being lost or severely damaged as a result of the defection. A DIP can usually still penetrate host cells, but requires another fully functional virus particle to co-infect a cell with it, in order to provide the lost factors.
A satellite is a subviral agent that depends on the coinfection of a host cell with a helper virus for its replication. Satellites can be divided into two major classes: satellite viruses and satellite nucleic acids. Satellite viruses, which are most commonly associated with plants, but are also found in mammals, arthropods, and bacteria, encode structural proteins to enclose their genetic material, which are therefore distinct from the structural proteins of their helper viruses. Satellite nucleic acids, in contrast, do not encode their own structural proteins, but instead are encapsulated by proteins encoded by their helper viruses. The genomes of satellites range upward from 359 nucleotides in length for satellite tobacco ringspot virus RNA (STobRV).

Tobamovirus is a genus of positive-strand RNA viruses in the family Virgaviridae. Many plants, including tobacco, potato, tomato, and squash, serve as natural hosts. Diseases associated with this genus include: necrotic lesions on leaves. The name Tobamovirus comes from the host and symptoms of the first virus discovered.
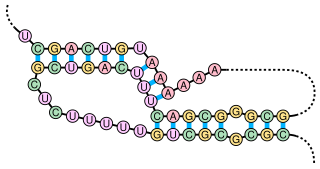
A pseudoknot is a nucleic acid secondary structure containing at least two stem-loop structures in which half of one stem is intercalated between the two halves of another stem. The pseudoknot was first recognized in the turnip yellow mosaic virus in 1982. Pseudoknots fold into knot-shaped three-dimensional conformations but are not true topological knots.

Potyvirus is a genus of positive-strand RNA viruses in the family Potyviridae. Plants serve as natural hosts. The genus is named after member virus potato virus Y. Potyviruses account for ~30% of the currently known plant viruses. Like begomoviruses, members of this genus may cause significant losses in agricultural, pastoral, horticultural and ornamental crops. More than 200 species of aphids spread potyviruses and most are from the subfamily Aphidinae. The genus contains 190 species.

Closterovirus, also known as beet yellows viral group, is a genus of viruses, in the family Closteroviridae. Plants serve as natural hosts. There are 17 species in this genus. Diseases associated with this genus include: yellowing and necrosis, particularly affecting the phloem. This genus has a probably worldwide distribution and includes among other viral species the Beet yellows virus and Citrus tristeza virus, rather economically important plant diseases. At least some species require vectors such as aphids or mealybugs for their transmission from plant to plant.

Crinivirus, formerly the lettuce infectious yellows virus group, is a genus of viruses, in the family Closteroviridae. They are linear, single-stranded positive sense RNA viruses. There are 14 species in this genus. Diseases associated with this genus include: yellowing and necrosis, particularly affecting the phloem.
Luteovirus is a genus of viruses, in the family Tombusviridae. There are 13 species in this genus. Plants serve as natural hosts. The geographical distribution of Luteoviruses is widespread, with the virus primarily infecting plants via transmission by aphid vectors. The virus only replicates within the host cell and not within the vector. The name 'luteovirus' arises from the Latin luteus, which is translated as 'yellow'. Luteovirus was given this name due to the symptomatic yellowing of the plant that occurs as a result of infection.

In molecular biology, the coronavirus frameshifting stimulation element is a conserved stem-loop of RNA found in coronaviruses that can promote ribosomal frameshifting. Such RNA molecules interact with a downstream region to form a pseudoknot structure; the region varies according to the virus but pseudoknot formation is known to stimulate frameshifting. In the classical situation, a sequence 32 nucleotides downstream of the stem is complementary to part of the loop. In other coronaviruses, however, another stem-loop structure around 150 nucleotides downstream can interact with members of this family to form kissing stem-loops and stimulate frameshifting.

The Coronavirus packaging signal is a conserved cis-regulatory element found in Betacoronavirus. It has an important role in regulating the packaging of the viral genome into the capsid. As part of the viral life cycle, within the infected cell, the viral genome becomes associated with viral proteins and assembles into new infective progeny viruses. This process is called packaging and is vital for viral replication.
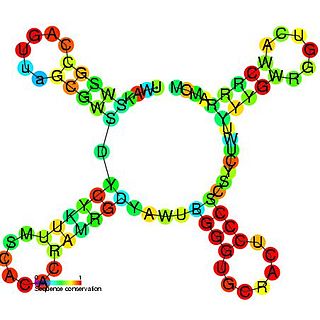
The tymoviruses/pomovirusesfamily tRNA-like 3' UTR element is an RNA element found in the 3' UTR of some viruses. This element acts in conjunction with UPSK RNA and a 5'-cap to enhance translation. The secondary structure of this RNA element is a cloverleaf that resembles tRNA.

Benyvirus is a genus of viruses, in the family Benyviridae. Plant serve as natural hosts. There are four species in this genus. Diseases associated with this genus include: BNYVV: rhizomania.

Sobemovirus is a genus of viruses. Plants serve as natural hosts. There are 20 species in this genus. Diseases associated with this genus include: mosaics and mottles.
Stockholm format is a multiple sequence alignment format used by Pfam and Rfam to disseminate protein and RNA sequence alignments. The alignment editors Ralee , Belvu and Jalview support Stockholm format as do the probabilistic database search tools, Infernal and HMMER, and the phylogenetic analysis tool Xrate. Stockholm format files often have the filename extension .sto or .stk.

Transfer RNA-like structures are RNA sequences, which have a similar tertiary structure to tRNA; they frequently contain a pseudoknot close to the 3' end. The presence of tRNA-like structures has been demonstrated in many plant virus RNA genomes. These tRNA-like structures are linked to regulation of plant virus replication.

Red clover necrotic mosaic virus (RCNMV) contains several structural elements present within the 3′ and 5′ untranslated regions (UTR) of the genome that enhance translation. In eukaryotes transcription is a prerequisite for translation. During transcription the pre-mRNA transcript is processes where a 5′ cap is attached onto mRNA and this 5′ cap allows for ribosome assembly onto the mRNA as it acts as a binding site for the eukaryotic initiation factor eIF4F. Once eIF4F is bound to the mRNA this protein complex interacts with the poly(A) binding protein which is present within the 3′ UTR and results in mRNA circularization. This multiprotein-mRNA complex then recruits the ribosome subunits and scans the mRNA until it reaches the start codon. Transcription of viral genomes differs from eukaryotes as viral genomes produce mRNA transcripts that lack a 5’ cap site. Despite lacking a cap site viral genes contain a structural element within the 5’ UTR known as an internal ribosome entry site (IRES). IRES is a structural element that recruits the 40s ribosome subunit to the mRNA within close proximity of the start codon.

In molecular biology, the Tymovirus coat protein refers to the protein coat of a virus order, named Tymovirales. More specifically this protein signature is found only in coat proteins from the related tymoviruses. The coat protein (CP) is also known as the virion protein. The virus coat is composed of 180 copies of the coat protein arranged in an icosahedral shell.
In molecular biology, Turnip crinkle virus (TCV) hairpin H4 is an RNA hairpin found at the 3' end of the Turnip crinkle virus (TCV) genome.
Coronavirus genomes are positive-sense single-stranded RNA molecules with an untranslated region (UTR) at the 5′ end which is called the 5′ UTR. The 5′ UTR is responsible for important biological functions, such as viral replication, transcription and packaging. The 5′ UTR has a conserved RNA secondary structure but different Coronavirus genera have different structural features described below.
