
Pro-opiomelanocortin (POMC) is a precursor polypeptide with 241 amino acid residues. POMC is synthesized in corticotrophs of the anterior pituitary from the 267-amino-acid-long polypeptide precursor pre-pro-opiomelanocortin (pre-POMC), by the removal of a 26-amino-acid-long signal peptide sequence during translation. POMC is part of the central melanocortin system.

Leptin is a protein hormone predominantly made by adipose cells and its primary role is likely to regulate long-term energy balance.

Orexin, also known as hypocretin, is a neuropeptide that regulates arousal, wakefulness, and appetite. The most common form of narcolepsy, type 1, in which the individual experiences brief losses of muscle tone, is caused by a lack of orexin in the brain due to destruction of the cells that produce it. It exists in the forms of orexin-A and orexin-B.
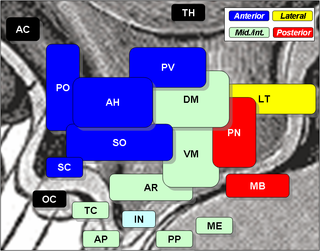
The arcuate nucleus of the hypothalamus is an aggregation of neurons in the mediobasal hypothalamus, adjacent to the third ventricle and the median eminence. The arcuate nucleus includes several important and diverse populations of neurons that help mediate different neuroendocrine and physiological functions, including neuroendocrine neurons, centrally projecting neurons, and astrocytes. The populations of neurons found in the arcuate nucleus are based on the hormones they secrete or interact with and are responsible for hypothalamic function, such as regulating hormones released from the pituitary gland or secreting their own hormones. Neurons in this region are also responsible for integrating information and providing inputs to other nuclei in the hypothalamus or inputs to areas outside this region of the brain. These neurons, generated from the ventral part of the periventricular epithelium during embryonic development, locate dorsally in the hypothalamus, becoming part of the ventromedial hypothalamic region. The function of the arcuate nucleus relies on its diversity of neurons, but its central role is involved in homeostasis. The arcuate nucleus provides many physiological roles involved in feeding, metabolism, fertility, and cardiovascular regulation.

Ghrelin is a hormone produced by enteroendocrine cells of the gastrointestinal tract, especially the stomach, and is often called a "hunger hormone" because it increases the drive to eat. Blood levels of ghrelin are highest before meals when hungry, returning to lower levels after mealtimes. Ghrelin may help prepare for food intake by increasing gastric motility and stimulating the secretion of gastric acid.

Neuropeptide Y (NPY) is a 36 amino-acid neuropeptide that is involved in various physiological and homeostatic processes in both the central and peripheral nervous systems. It is secreted alongside other neurotransmitters such as GABA and glutamate.
The melanocortins are a family of neuropeptide hormones which are the ligands of the melanocortin receptors The melanocortin system consists of melanocortin receptors, ligands, and accessory proteins. The genes of the melanocortin system are found in chordates. Melanocortins were originally named so because their earliest known function was in melanogenesis. It is now known that the melanocortin system regulates diverse functions throughout the body, including inflammatory response, fibrosis, melanogenesis, steroidogenesis, energy homeostasis, sexual function, and exocrine gland function.

Agouti-signaling protein is a protein that in humans is encoded by the ASIP gene. It is responsible for the distribution of melanin pigment in mammals. Agouti interacts with the melanocortin 1 receptor to determine whether the melanocyte produces phaeomelanin, or eumelanin. This interaction is responsible for making distinct light and dark bands in the hairs of animals such as the agouti, which the gene is named after. In other species such as horses, agouti signalling is responsible for determining which parts of the body will be red or black. Mice with wildtype agouti will be grey, with each hair being partly yellow and partly black. Loss of function mutations in mice and other species cause black fur coloration, while mutations causing expression throughout the whole body in mice cause yellow fur and obesity.

Peptide YY (PYY) also known as peptide tyrosine tyrosine is a peptide that in humans is encoded by the PYY gene. Peptide YY is a short peptide released from cells in the ileum and colon in response to feeding. In the blood, gut, and other elements of periphery, PYY acts to reduce appetite; similarly, when injected directly into the central nervous system, PYY is also anorexigenic, i.e., it reduces appetite.
Nesfatin-1 is a neuropeptide produced in the hypothalamus of mammals. It participates in the regulation of hunger and fat storage. Increased nesfatin-1 in the hypothalamus contributes to diminished hunger, a 'sense of fullness', and a potential loss of body fat and weight.
Growth hormone secretagogue receptor(GHS-R), also known as ghrelin receptor, is a G protein-coupled receptor that binds growth hormone secretagogues (GHSs), such as ghrelin, the "hunger hormone". The role of GHS-R is thought to be in regulating energy homeostasis and body weight. In the brain, they are most highly expressed in the hypothalamus, specifically the ventromedial nucleus and arcuate nucleus. GSH-Rs are also expressed in other areas of the brain, including the ventral tegmental area, hippocampus, and substantia nigra. Outside the central nervous system, too, GSH-Rs are also found in the liver, in skeletal muscle, and even in the heart.

The KiSS1-derived peptide receptor is a G protein-coupled receptor which binds the peptide hormone kisspeptin (metastin). Kisspeptin is encoded by the metastasis suppressor gene KISS1, which is expressed in a variety of endocrine and gonadal tissues. Activation of the kisspeptin receptor is linked to the phospholipase C and inositol trisphosphate second messenger cascades inside the cell.

Melanocortin 4 receptor (MC4R) is a melanocortin receptor that in humans is encoded by the MC4R gene. It encodes the MC4R protein, a G protein-coupled receptor (GPCR) that binds α-melanocyte stimulating hormone (α-MSH). In mouse models, MC4 receptors have been found to be involved in feeding behaviour, the regulation of metabolism, sexual behaviour, and male erectile function.
Melanocortin 3 receptor (MC3R) is a protein that in humans is encoded by the MC3R gene.

Pancreatic polypeptide receptor 1, also known as Neuropeptide Y receptor type 4, is a protein that in humans is encoded by the PPYR1 gene.
The central melanocortin system is defined anatomically as a collection of central nervous system circuits which include:
Hunger is a sensation that motivates the consumption of food. The sensation of hunger typically manifests after only a few hours without eating and is generally considered to be unpleasant. Satiety occurs between 5 and 20 minutes after eating. There are several theories about how the feeling of hunger arises. The desire to eat food, or appetite, is another sensation experienced with regards to eating.

Teleost leptins are a family of peptide hormones found in fish (teleostei) that are orthologs of the mammalian hormone leptin. The teleost and mammalian leptins appear to have similar functions, namely, regulation of energy intake and expenditure.

Melanocortin 2 receptor accessory protein 2 is a protein that in humans is encoded by the MRAP2 gene. MRAP2 is a transmembrane accessory protein to a family of five receptors called the melanocortin receptors (MC1-5). Human genome sequencing analysis led to the discovery of MRAP2, Human MRAP2 gene is located in chromosome 6q14.3, a different chromosomal location from that of human MRAP which is a paralogy to MRAP2. MRAP2 is thought to be involved in regulating the expression of the melanocortin (MC1-5) and some non-melanocortin receptors such as ghrelin receptor (GHSR-1a), orexin (OX1R) receptor and prokineticin receptor (PKR-1).
Pathophysiology of obesity is the study of disordered physiological processes that cause, result from, or are otherwise associated with obesity. A number of possible pathophysiological mechanisms have been identified which may contribute in the development and maintenance of obesity.







