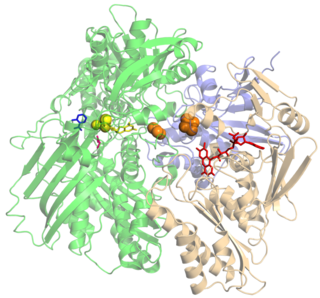
an aldehyde + H2O + 2 oxidized ferredoxin ⇌ an acid + 3 H+ + 2 reduced ferredoxin
This enzyme belongs to the family of oxidoreductases, specifically those acting on the aldehyde or oxo group of donor with an iron-sulfur protein as acceptor. The systematic name of this enzyme class is aldehyde:ferredoxin oxidoreductase. This enzyme is also called AOR. It is a relatively rare example of a tungsten-containing protein.[1]
Occurrence
The active site of the AOR family feature an oxo-tungsten center bound to a pair of molybdopterincofactors (which does not contain molybdenum) and an 4Fe-4S cluster.[2][3] This family includes AOR, formaldehyde ferredoxin oxidoreductase (FOR), glyceraldehyde-3-phosphate ferredoxin oxidoreductase (GAPOR), all isolated from hyperthermophilic archea;[2]carboxylic acidreductase found in clostridia;[4] and hydroxycarboxylate viologen oxidoreductase from Proteus vulgaris, the sole member of the AOR family containing molybdenum.[5] GAPOR may be involved in glycolysis,[6] but the functions of the other proteins are not yet clear. AOR has been proposed to be the primary enzyme responsible for oxidising the aldehydes that are produced by the 2-keto acid oxidoreductases.[7]
AOR is found in hyperthermophillic archaea, Pyrococcus furiosus.[1] The archaeons Pyrococcus ES-4 strain and Thermococcus ES-1 strain differ by their substrate specificity: AFOs show a broader size range of its aldehyde substrates. Its primary role is to oxidize aldehyde coming derived from the metabolism of amino acids and glucoses.[8] Aldehyde Ferredoxin Oxidoreductase is a member of an AOR family, which includes glyceraldehyde-3-phosphate ferredoxin oxidoreductase (GAPOR) and Formaldehyde Ferredoxin Oxidoreductase.[3]
Function
AOR functions at high temperature conditions (~80 degrees Celsius) at an optimal pH of 8-9. It is oxygen-sensitive as it loses bulk of its activity from oxygen exposure and works in the cytoplasm where it is a reducing environment. Thus, either exposure to oxygen or lowering of the temperature causes an irreversible loss of its catalytic properties. Also, as a result of oxygen sensitivity of AOR, purification of the enzyme is done under anoxic environments.[8]
It is proposed that AOR has a role in the Entner-Doudoroff pathway (glucose degradation) due to its increased activity with maltose incorporation.[3] However, other proposals include its role in oxidation of amino acid metabolism aldehyde side products coming from de-aminated 2-ketoacids. The main substrates for aldehyde ferredoxin oxidoreductase are acetaldehyde, phenylacetaldehyde, and isovalerdehyde, which is a metabolic product from common amino acids and glucose.[8] For example, acetaldehyde reaches its kcat/KM value up to 22.0 μM-1s-1.[8] In fact, some microorganisms only make use of amino acids as a carbon source, such as Thermococcus strain ES1; thus, they utilize aldehyde ferredoxin oxidoreductase to metabolize the amino acid carbon source.[8]
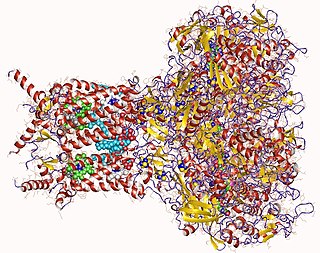
AOR is homodimeric. Each 67kDa subunit contains 1 tungsten and 4-5 Iron atoms.[3] The two subunits are bridged by a low spin Iron center. It is believed that the two subunits function independently.[3]
Tungsten-pterin
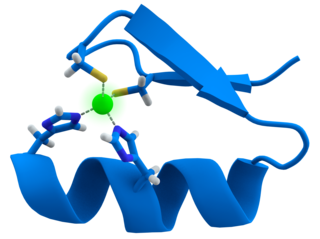
Tungsten in the active site of AOR adopts a distorted square pyramidal geometry bound an oxo/hydroxo ligand and the dithiolene substituents of two molybdopterin cofactors.[3]
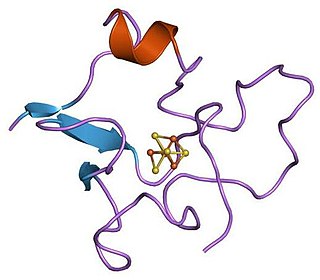
Molybdopterin cofactor, shown in the dithiol protonation state.
Two molybdopterin cofactors bind tungsten,[9] as observed in many related enzymes.[9] Tungsten is not bonded directly to the protein.[9] Phosphate centers pendant on the cofactor are bound to a Mg2+, which is also bound by Asn93 and Ala183 to complete its octahedral coordination sphere.[3][9] Thus, pterin and Tungsten atoms are connected to the AOR enzyme primarily through pterin's Hydrogen bonding networks with the amino acid residues.[3] In addition, two water ligands that occupy the octahedral geometry take part in hydrogen bonding networks with pterin, phosphate, and Mg2+.[9] While [Fe4S4] cluster is bound by four Cys ligands, Pterin - rich in amino and ether linkages - interacts with the Asp-X-X-Gly-Leu-(Cys/Asp) sequences in the AOR enzyme.[3] In such sequence, Cys494 residue is also hydrogen bonded to the [Fe4S4] cluster.[3] This indicates that Cys494 residue connects the Tungsten site and the [Fe4S4] cluster site in the enzyme.[3] Iron atom in the cluster is additionally bound by three other Cystein ligands: .[9] Also, another linker amino acid residue between ferredoxin cluster and pterin is the Arg76, which hydrogen bonds to both pterin and ferredoxin.[3] It is proposed that such hydrogen bonding interactions imply pterin cyclic ring system as an electron carrier.[3] Additionally the C=O center of the pterin binds Na+.[8] The W=O center is proposed, not verified crystallographically.[9]
AOR consists of three domains, domain 1, 2, and 3.[8] While domain 1 contains pterin bound to tungsten, the other two domains provide a channel from tungsten to protein's surface (15 Angstroms in length) in order to allow specific substrates to enter the enzyme through its channel.[8] In the active site, this pterin molecules is in a saddle-like conformation (500 to the normal plane) to “sit” on the domain 1 which also takes on a form with beta sheets to accommodate the Tungsten-Pterin site.[8]
Iron
The iron center in between the two subunits serve a structural role in AOR.[8] Iron metal atoms takes on a tetrahedral conformation while the ligand coordination comes from two histidines and glutamic acids.[8] This is not known to have any functional role in the redox activity of the protein.[8]
Fe4S4 centre
[Fe4S4] cluster in AOR is different in some aspects to other ferredoxin molecules.[3] EPR measurements confirm that it serves as a one-electron shuttle.[3]
Aldehyde ferredoxin oxidoreductase mechanism
In the catalytic cycle, W(VI) (tungsten "six") converts to W(IV) concomitant with oxidation of the aldehyde to a carboxylic acid (equivalently, a carboxylate).[3] A W(V) intermediate can be detected by EPR spectroscopy.[3][8]
The redox equivalents are provided by the 4Fe-4S cluster.
A tyrosine residue is proposed to activate the electrophilic centre of aldehydes by H-bonding to the carbonyl oxygen atom, coordinated to the W centre.[10] A glutamic acid residue near the active site activates a water molecule for a nucleophilic attack on aldehyde carbonyl center.[10] After nucleophilic attack by water, hydride is transferred to oxo-tungsten sie thus, .[10] Subsequently, W(VI) is regenerated by electron transfer to the 4Fe-4S center.[10] With formaldehyde ferredoxin oxidoreductase, Glu308 and Tyr 416 would be involved while Glu313 and His448 is shown to be present in AOR active site.[9][10]
Related Research Articles
Xanthine oxidase is a form of xanthine oxidoreductase, a type of enzyme that generates reactive oxygen species. These enzymes catalyze the oxidation of hypoxanthine to xanthine and can further catalyze the oxidation of xanthine to uric acid. These enzymes play an important role in the catabolism of purines in some species, including humans.
A cofactor is a non-protein chemical compound or metallic ion that is required for an enzyme's role as a catalyst. Cofactors can be considered "helper molecules" that assist in biochemical transformations. The rates at which these happen are characterized in an area of study called enzyme kinetics. Cofactors typically differ from ligands in that they often derive their function by remaining bound.
Ferredoxins are iron–sulfur proteins that mediate electron transfer in a range of metabolic reactions. The term "ferredoxin" was coined by D.C. Wharton of the DuPont Co. and applied to the "iron protein" first purified in 1962 by Mortenson, Valentine, and Carnahan from the anaerobic bacterium Clostridium pasteurianum.
Pterin is a heterocyclic compound composed of a pteridine ring system, with a "keto group" and an amino group on positions 4 and 2 respectively. It is structurally related to the parent bicyclic heterocycle called pteridine. Pterins, as a group, are compounds related to pterin with additional substituents. Pterin itself is of no biological significance.
Pyrococcus furiosus is a heterotrophic, strictly anaerobic, extremophilic, model species of archaea. It is classified as a hyperthermophile because it thrives best under extremely high temperatures, and is notable for having an optimum growth temperature of 100 °C. P. furiosus belongs to the Pyrococcus genus, most commonly found in extreme environmental conditions of hydrothermal vents. It is one of the few prokaryotic organisms that has enzymes containing tungsten, an element rarely found in biological molecules.
DMSO reductase is a molybdenum-containing enzyme that catalyzes reduction of dimethyl sulfoxide (DMSO) to dimethyl sulfide (DMS). This enzyme serves as the terminal reductase under anaerobic conditions in some bacteria, with DMSO being the terminal electron acceptor. During the course of the reaction, the oxygen atom in DMSO is transferred to molybdenum, and then reduced to water.
Molybdopterins are a class of cofactors found in most molybdenum-containing and all tungsten-containing enzymes. Synonyms for molybdopterin are: MPT and pyranopterin-dithiolate. The nomenclature for this biomolecule can be confusing: Molybdopterin itself contains no molybdenum; rather, this is the name of the ligand that will bind the active metal. After molybdopterin is eventually complexed with molybdenum, the complete ligand is usually called molybdenum cofactor.
Formate dehydrogenases are a set of enzymes that catalyse the oxidation of formate to carbon dioxide, donating the electrons to a second substrate, such as NAD+ in formate:NAD+ oxidoreductase (EC 1.17.1.9) or to a cytochrome in formate:ferricytochrome-b1 oxidoreductase (EC 1.2.2.1). This family of enzymes has attracted attention as inspiration or guidance on methods for the carbon dioxide fixation, relevant to global warming.
In enzymology, a 2-oxoglutarate synthase (EC 1.2.7.3) is an enzyme that catalyzes the chemical reaction
In enzymology, a 3-methyl-2-oxobutanoate dehydrogenase (ferredoxin) (EC 1.2.7.7) is an enzyme that catalyzes the chemical reaction
In enzymology, a glyceraldehyde-3-phosphate dehydrogenase (ferredoxin) (EC 1.2.7.6) is an enzyme that catalyzes the chemical reaction
In enzymology, an indolepyruvate ferredoxin oxidoreductase (EC 1.2.7.8) is an enzyme that catalyzes the chemical reaction
In enzymology, an ethylbenzene hydroxylase (EC 1.17.99.2) is an enzyme that catalyzes the chemical reaction
High potential iron-sulfur proteins (HIPIP) are a class of iron-sulfur proteins. They are ferredoxins that participate in electron transfer in photosynthetic bacteria as well as in Paracoccus denitrificans.
Molybdopterin synthase (EC 2.8.1.12, MPT synthase) is an enzyme required to synthesize molybdopterin (MPT) from precursor Z (now known as cyclic pyranopterin monophosphate). Molydopterin is subsequently complexed with molybdenum to form molybdenum cofactor (MoCo). MPT synthase catalyses the following chemical reaction:
A transition metal oxo complex is a coordination complex containing an oxo ligand. Formally O2-, an oxo ligand can be bound to one or more metal centers, i.e. it can exist as a terminal or (most commonly) as bridging ligands (Fig. 1). Oxo ligands stabilize high oxidation states of a metal. They are also found in several metalloproteins, for example in molybdenum cofactors and in many iron-containing enzymes. One of the earliest synthetic compounds to incorporate an oxo ligand is potassium ferrate (K2FeO4), which was likely prepared by Georg E. Stahl in 1702.
Hydrogenase (NAD+, ferredoxin) (EC 1.12.1.4, bifurcating [FeFe] hydrogenase) is an enzyme with systematic name hydrogen:NAD+, ferredoxin oxidoreductase. This enzyme catalyses the following chemical reaction
Transition metal thiolate complexes are metal complexes containing thiolate ligands. Thiolates are ligands that can be classified as soft Lewis bases. Therefore, thiolate ligands coordinate most strongly to metals that behave as soft Lewis acids as opposed to those that behave as hard Lewis acids. Most complexes contain other ligands in addition to thiolate, but many homoleptic complexes are known with only thiolate ligands. The amino acid cysteine has a thiol functional group, consequently many cofactors in proteins and enzymes feature cysteinate-metal cofactors.
Evolution of metal ions in biological systems refers to the incorporation of metallic ions into living organisms and how it has changed over time. Metal ions have been associated with biological systems for billions of years, but only in the last century have scientists began to truly appreciate the scale of their influence. Major and minor metal ions have become aligned with living organisms through the interplay of biogeochemical weathering and metabolic pathways involving the products of that weathering. The associated complexes have evolved over time.
In enzymology, a formylmethanofuran dehydrogenase (EC 1.2.99.5) is an enzyme that catalyzes the chemical reaction:
References
1 2 Majumdar A, Sarkar S (May 2011). "Bioinorganic chemistry of molybdenum and tungsten enzymes: A structural–functional modeling approach". Coordination Chemistry Reviews. 255 (9–10): 1039–1054. doi:10.1016/j.ccr.2010.11.027.
↑ White H, Strobl G, Feicht R, Simon H (September 1989). "Carboxylic acid reductase: a new tungsten enzyme catalyses the reduction of non-activated carboxylic acids to aldehydes". Eur. J. Biochem. 184 (1): 89–96. doi:10.1111/j.1432-1033.1989.tb14993.x. PMID2550230.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.