Scholarly interest in the process and functions of dreaming has been present since Sigmund Freud's interpretations in the 1900s. The neurology of dreaming has remained misunderstood until recent distinctions, however. The information available via modern techniques of brain imaging has provided new bases for the study of the dreaming brain. The bounds that such technology has afforded has created an understanding of dreaming that seems ever-changing; even now questions still remain as to the function and content of dreams.
Preliminary observations into the neurology of dreaming were reported in 1951 by George Humphrey and Oliver Zangwill. Their report noted two cases of brain injury that resulted in the complete or almost complete cessation of dreaming. Both patients had undergone damage to posterior parietal regions, one of which involved predominately the left side of the parieto-occipital areas. Additional effects involved hemianopia, reduced visualization (in waking state), and disturbances in visual memory. Patients reported that their visual images were dim and hard to evoke. Although they reported only two cases, Humphrey and Zangwill offered preliminary ideas about neurological components of dreaming, specifically the association of forebrain areas and the link between visual imaging and the ability to dream.[1]
Methodological issues in scientific dream studies
There are several difficulties encountered while studying subjective experiences like dreaming. Methodologies in dream studies are abound with conceptual complexities and limitations.
Reliance on verbal reports
One significant shortcoming of dream studies is the necessary reliance on verbal reports. The dream event is reduced to a verbal report which is only an account of the subject's memory of the dream, not the subject's experience of the dream itself. These verbal reports are also at risk of being influenced by a number of factors. First, dreams involve multiple pseudo-sensory, emotional and motoric elements. The dream report is only narrative, which makes capturing the whole picture difficult. Verbal reports face other difficulties like forgetting. Dreams and reports of dreams are produced in distinct states of consciousness resulting in a delay between the dream event and its recall while awake. During this time lag forgetting may occur resulting in an incomplete report. Forgetting is proportional to the amount of time elapsed between the experience and its recall.[2] Also, remembering is exposed to interference at the recall stage and some information is not accessible to recall.[2] Reconstructing the dream from memory while awake might affect the accuracy of recall because the subject may report more information than actually experienced, and sequence of events may be reordered.[2] Another issue is the difficulty of verbally describing mostly visual subjective experiences like those found in dreams (e.g. unreal objects, bizarre experiences, emotions). Furthermore, subjects may intentionally fail to report embarrassing, immoral, or private dream experiences for fear of judgement, which results in censored, incomplete reports.
The sleep laboratory environment
The sleep laboratory environment is another major source of methodological issues. Sleep laboratories are an unnatural, awkward environment for sleeping. The subject may feel discomfort and anxiety, which may make sleep more difficult and of inferior quality. This is the well-known first night effect. Complete adaptation to the sleep laboratory may take four days or longer,[3] which is longer than the duration of most laboratory studies. Also, the content of dreams at the laboratory has been observed to be different from dreams at home.[3] Similarly, the laboratory environment may alter the content of dreams recalled from spontaneous awakenings at the end of a night's sleep, as indicated by high frequency of laboratory references in morning spontaneous awakenings in REM and NREM dream reports.[3]
Statistical concerns
Statistical concerns in dream studies are another cause of methodological issues. Many investigators used small samples for sleep studies and statistical parametric mapping (a technique for examining differences in brain activity recorded during functional neuroimaging experiments).[4] Results obtained from small samples must be interpreted with caution due to inherent statistical problems associated with small samples.
Technological limitations
Technological limitations also pose methodological problems. Measures of global brain activity like electroencephalogram (EEG) voltage averaging or cerebral blood flow cannot identify small but influential neuronal populations like the locus coeruleus, the raphe nucleus and the pedunculopontine tegmental nucleus, which reveal mechanistic and functional details in dreaming.[5] Despite these shortcomings, it is widely agreed that clinical findings and data obtained from neuro-imaging are valid, affirming neuro-imaging as an essential tool in cognitive neuroscience.
Lesion and activation interpretations
Brain-damaged patients offer valuable but rare information about human brain mechanisms. Eugene Aserinsky and Nathaniel Kleitman observed REM sleep and concluded that it was the physiological manifestation of dreaming. This was assumed to be a breakthrough in the understanding of such an elusive process as dreaming. Indeed, 95% of subjects awakened during REM reported that they had been dreaming whereas only about 5-10% reported dreams after being awakened during non-REM sleep (NREM).[6]
REM and NREM dream reports compared
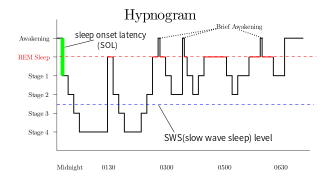
The REM portion of sleep is outlined in red
There are several important differences between REM and NREM dream reports. There is disagreement amongst experts about the existence of qualitative differences, but there is a general consensus that there are quantitative differences.[citation needed]
It has been recognized that following REM awakenings dream reports are obtained substantially more frequently than after NREM awakenings.[7] Subjects dream reports are related to the length of REM sleep. Word count and subjectively estimated dream duration increase as length of preceding REM sleep increases, revealing a positive relationship.[8] Reports from REM awakenings tend to be longer, more multimodal perceptually, have intensified emotionality, and are less reminiscent of waking life than NREM awakenings.[5] Judges are able to differentiate unaltered REM and NREM dream reports, while some subjects are able to discern whether they themselves had been awakened from REM or NREM.[5]
The characteristics of REM sleep consistently contain a similar set of features. While dreaming, people regularly falsely believe that they are awake unless they implement lucidity. Dreams contain multimodal pseudo-perceptions; sometimes any or all sensory modalities are present, but most often visual and motoric.[9] Dream imagery can change quickly and is regularly of a bizarre nature, but reports also contain many images and events that are a part of day-to-day life.[9] In dreams there is a reduction or absence of self-reflection or other forms of meta-cognition relative to during waking life.[5] Dreams are also characterized by a lack of "orientational stability; persons, times, and places are fused, plastic, incongruous and discontinuous".[9] In addition, dreams form a single narrative to explain and integrate all dream elements.[9] Lastly, NREM reports contain thought-like mentation and depictions of current concerns more frequently than REM reports.[5]
Neuroanatomy of dreaming
REM sleep and dreaming
Aserinsky and Kleitman's discovery prompted further research into the brain mechanism involved in REM sleep (and by their assumption, dreaming). It was found that REM is generated by a small region of cells located in the brain stem called the pons (it sits slightly above the spinal cord at the nape of the neck). The pons releases acetylcholine which travels to parts of the forebrain. Cholinergic activation of these higher areas was thought to result in the meaningless images that make up our dreams. This process is switched off by noradrenaline and serotonin which are also released by the brain stem.
The formation of the Activation-Synthesis Model put forth by Allan Hobson and McCarley in 1975 rested largely on these discoveries. Their model posits that dreams are actively generated by the brain stem and then passively synthesized by the forebrain. That is, the cholinergic activation that occurs in any forebrain areas (via transmission from brain stem) results in attempts by the brain's cognitive areas to enforce sense or structure onto meaningless activation.[10] Cerebral areas were not thought to play any sort of causal role because REM sleep occurs as long as the pons is intact, even if higher areas are disconnected or removed;[6] an inference based on the assumption that REM sleep is dreaming.
A shift to NREM
Although this assumption has remained a predominant view, disputing evidence has been present since the 60s. Foulkes for example reported that complex mentation is indeed possible during NREM. Previously participants reported dreams mainly after being awakened from REM, however upon awakening during NREM Foulkes asked subjects about what had just been passing through their heads, rather than whether or not they were dreaming. As many as 50% of subjects reported some form of complex mentation.[6] Furthermore, these NREM dreams seemed to cluster around specific sleep stages (stage 1 and late stages).[10] This offered evidence that dreaming was not restricted nor caused by mechanisms controlling REM sleep, and that perhaps there are entirely different brain areas associated with dreaming.
An investigation of the differential brain structures can be conducted by clinico-anatomical correlations. Here, the mechanisms associated with REM sleep are removed to observe whether there is a cessation in dreaming as well, then the areas thought to be associated with dreaming are removed to see if REM sleep is also made impossible.[6] These studies, with the exception of natural accidents, are conducted with animals. A main problem with obliterating REM sleep is that the associated area, the brain stem, is responsible for consciousness. Lesions large enough to stop REM completely can also render the subject unconscious.[10] Supporting evidence did come from the flip-side of clinico-anatomical correlations however. In a compilation of all reported cases of dream cessation (111 cases in all) damage was located in an entirely different area of the brain than the brain stem.[10] Furthermore, REM sleep was maintained. Remember that the pons is crucial for REM. Loss of dreaming only occurred when higher parts of the cerebral hemispheres were damaged. REM sleep is controlled by cholinergic activation in the pons. It is now believed that dreaming may be a dopaminergic process that occurs in limbic and frontal areas of the brain.
Dopaminergic activation
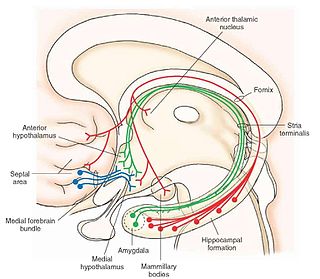
Two main frontal areas have been implicated in the dream process. The first involves the deep white matter of the frontal lobes (just above the eyes). The main systems at work here involve the mesolimbic and mesocorticaldopaminergic pathways. There are connecting fibres that run between frontal and limbic structures. A dopaminergic pathway runs from the ventral tegmental area, ascends through the lateral hypothalamus, various basal forebrain areas (nucleus basalis, stria terminalis, shell of nucleus accumbens) and terminates in the amygdala, anterior cingulate gyrus and frontal cortex. Damage to the dopaminergic pathway results in a loss of dreaming. Furthermore, chemical stimulation of the pathway (with L-DOPA for example) increases the frequency and vividness of dreams without affecting REM sleep.[10] The mesolimbic and mesocortical pathways are considered the seeking areas or the motivational command centers of the brain. Damage not only results in the loss of dreams but also of motivated behaviour.[6] Transection or inhibition of the dopamine pathway also reduces some positive symptoms of schizophrenia, many of which have been likened to dream-like states. Drugs that block the system have anti-psychotic effects but also reduce excessive and vivid dreaming.[10] Further evidence that dreaming can occur independently of REM sleep is found in the occurrence of nocturnal seizures during NREM that often present themselves as nightmares. Activation here is seen in the temporal lobe, again a forebrain area.[6][10]
The evidence of the involvement of mesolimbic and mesocortical dopaminergic pathways suggests that dreaming occurs when a motivational component is activated. Only when this pathway is removed do dreams cease to occur. This system can be activated by mechanisms of REM sleep but can also occur independently during NREM stages of sleep.
Perceptual processing
Another area thought to be involved in the generation of dreams is the Parieto-Occipito-Temporal junction (PTO).[10] This is an area of grey cortex towards the back of the brain involved in the highest levels of perceptual processing. It is here that perceptions are converted into abstract thoughts and memories.[6] The PTO is also vital for mental imagery.[10] Damage specifically to this area results in complete loss of dreaming, however damage to lower levels of perceptual processing merely results in reduced aspects of dream imagery. This is the basis for the suggestion that dreaming involves a reversed sequence of perceptual events. Instead of bottom-up it is top-down (higher levels activating lower levels instead lower to higher). Activation of the motivational mechanisms in the brain would normally be directed toward goal-oriented actions. However, during sleep access to the motor system is blocked (by inactivation of the dorsolateral frontal convexity). As a result, activation moves backwards toward the perceptual areas. This is why the dreamer doesn't engage in motivated behaviours but imagines them. Furthermore, there is inactivation of the reflective system in the limbic brain which leads the dreamer to mistake the dream for reality. Damage to this area also results in the inability to distinguish dreams from reality during waking state.
Notes
↑ Humphrey, M. E., & Zangwill, O.L. (1951) Cessation of dreaming after brain injury. Journal of Neurology, Neurosurgery, and Psychiatry, 14, 322.
1 2 3 Schwartz, S., & Maquet, P. (2002). Sleep imaging and the neuro-psychological assessment of dreams. Trends in Cognitive Sciences, 6(1), 23-30.
1 2 3 Domhoff, B., & Kamiya, J. (1964). Problems in dream content study with objective indicators. Archives of General Psychiatry, 11(5) 519-532.
↑ Braun, A.R., Thomas, J., Nancy, J., Gwadry, W. F., Carson, R. E., Varga, M., Baldwin, P., Belenky, G., & Herscovitch, P. (1998) Dissociated Pattern of Activity in Visual Cortices and Their Projections During Human Rapid Eye Movement Sleep. Science 279, 91 – 95.
1 2 3 4 5 Hobson, J. A., Pace-Schott, E. F., & Stickgold, R. (2000). Dreaming and the brain: Toward a cognitive neuroscience of conscious states. Behavioral and Brain Sciences, 23(6), 793-842.
1 2 3 4 5 6 7 Solms, M. (2005). The interpretation of dreams and the neurosciences. Neuroscience and Freud's Dream Theory.
↑ Stoyva, J.M. (1965). Posthypnotically suggested dreams and the sleep cycle. Archives of General Psychiatry, 12(3), 287-294.
↑ (Dement, W., & Kleitman, N. (1957). The relation of eye movements during sleep to dream activity: An objective method for the study of dreaming. Journal of Experimental Psychology, 53(5), 339-346.)
1 2 3 4 Hobson, J.A. 1988. The dreaming brain: How the brain creates both the sense and the nonsense of dreams. Basic Books, NY.
1 2 3 4 5 6 7 8 9 Solms, M. (2000). Dreaming and REM sleep are controlled by different brain mechanisms. Behavioral and Brain Sciences, 23, 843-850.
Rapid eye movement sleep is a unique phase of sleep in mammals and birds, characterized by random rapid movement of the eyes, accompanied by low muscle tone throughout the body, and the propensity of the sleeper to dream vividly.
The mesolimbic pathway, sometimes referred to as the reward pathway, is a dopaminergic pathway in the brain. The pathway connects the ventral tegmental area in the midbrain to the ventral striatum of the basal ganglia in the forebrain. The ventral striatum includes the nucleus accumbens and the olfactory tubercle.
Physiological psychology is a subdivision of behavioral neuroscience that studies the neural mechanisms of perception and behavior through direct manipulation of the brains of nonhuman animal subjects in controlled experiments. This field of psychology takes an empirical and practical approach when studying the brain and human behavior. Most scientists in this field believe that the mind is a phenomenon that stems from the nervous system. By studying and gaining knowledge about the mechanisms of the nervous system, physiological psychologists can uncover many truths about human behavior. Unlike other subdivisions within biological psychology, the main focus of psychological research is the development of theories that describe brain-behavior relationships.
The sleep cycle is an oscillation between the slow-wave and REM (paradoxical) phases of sleep. It is sometimes called the ultradian sleep cycle, sleep–dream cycle, or REM-NREM cycle, to distinguish it from the circadian alternation between sleep and wakefulness. In humans, this cycle takes 70 to 110 minutes. Within the sleep of adults and infants there are cyclic fluctuations between quiet and active sleep. These fluctuations may persist during wakefulness as rest-activity cycles but are less easily discerned.
Dopaminergic pathways in the human brain are involved in both physiological and behavioral processes including movement, cognition, executive functions, reward, motivation, and neuroendocrine control. Each pathway is a set of projection neurons, consisting of individual dopaminergic neurons.
Non-rapid eye movement sleep (NREM), also known as quiescent sleep, is, collectively, sleep stages 1–3, previously known as stages 1–4. Rapid eye movement sleep (REM) is not included. There are distinct electroencephalographic and other characteristics seen in each stage. Unlike REM sleep, there is usually little or no eye movement during these stages. Dreaming occurs during both sleep states, and muscles are not paralyzed as in REM sleep. People who do not go through the sleeping stages properly get stuck in NREM sleep, and because muscles are not paralyzed a person may be able to sleepwalk. According to studies, the mental activity that takes place during NREM sleep is believed to be thought-like, whereas REM sleep includes hallucinatory and bizarre content. NREM sleep is characteristic of dreamer-initiated friendliness, compared to REM sleep where it is more aggressive, implying that NREM is in charge of simulating friendly interactions. The mental activity that occurs in NREM and REM sleep is a result of two different mind generators, which also explains the difference in mental activity. In addition, there is a parasympathetic dominance during NREM. The reported differences between the REM and NREM activity are believed to arise from differences in the memory stages that occur during the two types of sleep.
In the field of psychology, the subfield of oneirology is the scientific study of dreams. Current research seeks correlations between dreaming and current knowledge about the functions of the brain, as well as an understanding of how the brain works during dreaming as pertains to memory formation and mental disorders. The study of oneirology can be distinguished from dream interpretation in that the aim is to quantitatively study the process of dreams instead of analyzing the meaning behind them.
The reticular formation is a set of interconnected nuclei that are located in the brainstem, hypothalamus, and other regions. It is not anatomically well defined, because it includes neurons located in different parts of the brain. The neurons of the reticular formation make up a complex set of networks in the core of the brainstem that extend from the upper part of the midbrain to the lower part of the medulla oblongata. The reticular formation includes ascending pathways to the cortex in the ascending reticular activating system (ARAS) and descending pathways to the spinal cord via the reticulospinal tracts.
A K-complex is a waveform that may be seen on an electroencephalogram (EEG). It occurs during stage 2 NREM sleep. It is the "largest event in healthy human EEG". They are more frequent in the first sleep cycles.
Slow-wave sleep (SWS), often referred to as deep sleep, is the third stage of non-rapid eye movement sleep (NREM), where electroencephalography activity is characterised by slow delta waves.
The medial forebrain bundle (MFB) is a neural pathway containing fibers from the basal olfactory regions, the periamygdaloid region and the septal nuclei, as well as fibers from brainstem regions, including the ventral tegmental area and nigrostriatal pathway.
Hypnopompia is the state of consciousness leading out of sleep, a term coined by the psychical researcher Frederic Myers. Its mirror is the hypnagogic state at sleep onset; though often conflated, the two states are not identical and have a different phenomenological character. Hypnopompic and hypnagogic hallucinations are frequently accompanied by sleep paralysis, which is a state wherein one is consciously aware of one's surroundings but unable to move or speak.
Recurrent thalamo-cortical resonance or Thalamocortical oscillation is an observed phenomenon of oscillatory neural activity between the thalamus and various cortical regions of the brain. It is proposed by Rodolfo Llinas and others as a theory for the integration of sensory information into the whole of perception in the brain. Thalamocortical oscillation is proposed to be a mechanism of synchronization between different cortical regions of the brain, a process known as temporal binding. This is possible through the existence of thalamocortical networks, groupings of thalamic and cortical cells that exhibit oscillatory properties.
Robert W. McCarley, MD, (1937–2017) was Chair and Professor of Psychiatry at Harvard Medical School and the VA Boston Healthcare System. He is also Director of the Laboratory of Neuroscience located at the Brockton VA Medical Center and the McLean Hospital. McClarley was a prominent researcher in the field of sleep and dreaming as well as schizophrenia.
Ponto-geniculo-occipital waves or PGO waves are distinctive wave forms of propagating activity between three key brain regions: the pons, lateral geniculate nucleus, and occipital lobe; specifically, they are phasic field potentials. These waves can be recorded from any of these three structures during and immediately before REM sleep. The waves begin as electrical pulses from the pons, then move to the lateral geniculate nucleus residing in the thalamus, and end in the primary visual cortex of the occipital lobe. The appearances of these waves are most prominent in the period right before REM sleep, albeit they have been recorded during wakefulness as well. They are theorized to be intricately involved with eye movement of both wake and sleep cycles in many different animals.
Sleep onset is the transition from wakefulness into sleep. Sleep onset usually transits into non-rapid eye movement sleep but under certain circumstances it is possible to transit from wakefulness directly into rapid eye movement sleep.
The relationship between sleep and memory has been studied since at least the early 19th century. Memory, the cognitive process of storing and retrieving past experiences, learning and recognition, is a product of brain plasticity, the structural changes within synapses that create associations between stimuli. Stimuli are encoded within milliseconds; however, the long-term maintenance of memories can take additional minutes, days, or even years to fully consolidate and become a stable memory that is accessible. Therefore, the formation of a specific memory occurs rapidly, but the evolution of a memory is often an ongoing process.
Secondary consciousness is an individual's accessibility to their history and plans. The ability allows its possessors to go beyond the limits of the remembered present of primary consciousness. Primary consciousness can be defined as simple awareness that includes perception and emotion. As such, it is ascribed to most animals. By contrast, secondary consciousness depends on and includes such features as self-reflective awareness, abstract thinking, volition and metacognition. The term was coined by Gerald Edelman.
The activation-synthesis hypothesis, proposed by Harvard University psychiatrists John Allan Hobson and Robert McCarley, is a neurobiological theory of dreams first published in the American Journal of Psychiatry in December 1977. The differences in neuronal activity of the brainstem during waking and REM sleep were observed, and the hypothesis proposes that dreams result from brain activation during REM sleep. Since then, the hypothesis has undergone an evolution as technology and experimental equipment has become more precise. Currently, a three-dimensional model called AIM Model, described below, is used to determine the different states of the brain over the course of the day and night. The AIM Model introduces a new hypothesis that primary consciousness is an important building block on which secondary consciousness is constructed.
The neuroscience of sleep is the study of the neuroscientific and physiological basis of the nature of sleep and its functions. Traditionally, sleep has been studied as part of psychology and medicine. The study of sleep from a neuroscience perspective grew to prominence with advances in technology and the proliferation of neuroscience research from the second half of the twentieth century.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.