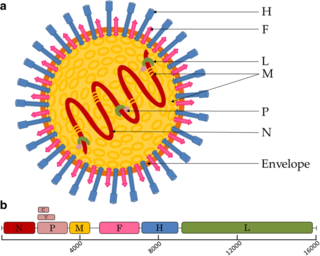
Paramyxoviridae is a family of negative-strand RNA viruses in the order Mononegavirales. Vertebrates serve as natural hosts. Diseases associated with this family include measles, mumps, and respiratory tract infections. The family has four subfamilies, 17 genera, three of which are unassigned to a subfamily, and 78 species.

Coronaviruses are a group of related RNA viruses that cause diseases in mammals and birds. In humans and birds, they cause respiratory tract infections that can range from mild to lethal. Mild illnesses in humans include some cases of the common cold, while more lethal varieties can cause SARS, MERS and COVID-19. In cows and pigs they cause diarrhea, while in mice they cause hepatitis and encephalomyelitis.

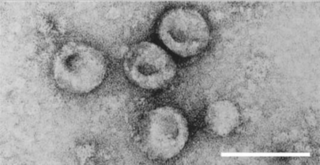
Severe-acute-respiratory-syndrome–related coronavirus is a species of virus consisting of many known strains. Two strains of the virus have caused outbreaks of severe respiratory diseases in humans: severe acute respiratory syndrome coronavirus 1, which caused the 2002–2004 outbreak of severe acute respiratory syndrome (SARS), and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), which is causing the ongoing pandemic of COVID-19. There are hundreds of other strains of SARSr-CoV, which are only known to infect non-human mammal species: bats are a major reservoir of many strains of SARSr-CoV; several strains have been identified in Himalayan palm civets, which were likely ancestors of SARS-CoV-1.
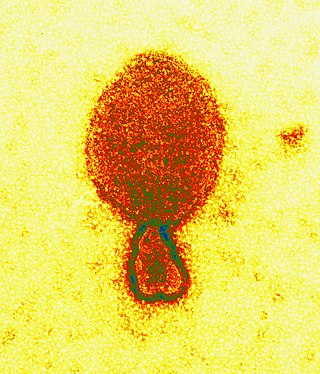
Henipavirus is a genus of negative-strand RNA viruses in the family Paramyxoviridae, order Mononegavirales containing six established species, and numerous others still under study. Henipaviruses are naturally harboured by several species of small mammals, notably pteropid fruit bats, microbats of several species, and shrews. Henipaviruses are characterised by long genomes and a wide host range. Their recent emergence as zoonotic pathogens capable of causing illness and death in domestic animals and humans is a cause of concern.

Measles morbillivirus(MeV), also called measles virus (MV), is a single-stranded, negative-sense, enveloped, non-segmented RNA virus of the genus Morbillivirus within the family Paramyxoviridae. It is the cause of measles. Humans are the natural hosts of the virus; no animal reservoirs are known to exist.
Rabies virus, scientific name Rabies lyssavirus, is a neurotropic virus that causes rabies in animals, including humans. Rabies transmission can occur through the saliva of animals and less commonly through contact with human saliva. Rabies lyssavirus, like many rhabdoviruses, has an extremely wide host range. In the wild it has been found infecting many mammalian species, while in the laboratory it has been found that birds can be infected, as well as cell cultures from mammals, birds, reptiles and insects. Rabies is reported in more than 150 countries and on all continents except Antarctica. The main burden of disease is reported in Asia and Africa, but some cases have been reported also in Europe in the past 10 years, especially in returning travellers.
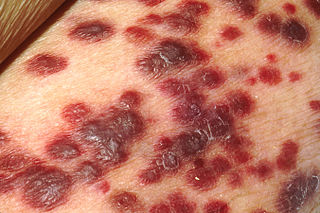
Kaposi's sarcoma-associated herpesvirus (KSHV) is the ninth known human herpesvirus; its formal name according to the International Committee on Taxonomy of Viruses (ICTV) is Human gammaherpesvirus 8, or HHV-8 in short. Like other herpesviruses, its informal names are used interchangeably with its formal ICTV name. This virus causes Kaposi's sarcoma, a cancer commonly occurring in AIDS patients, as well as primary effusion lymphoma, HHV-8-associated multicentric Castleman's disease and KSHV inflammatory cytokine syndrome. It is one of seven currently known human cancer viruses, or oncoviruses. Even after many years since the discovery of KSHV/HHV8, there is no known cure for KSHV associated tumorigenesis.

Nipah virus is a bat-borne, zoonotic virus that causes Nipah virus infection in humans and other animals, a disease with a very high mortality rate (40-75%). Numerous disease outbreaks caused by Nipah virus have occurred in South East Africa and Southeast Asia. Nipah virus belongs to the genus Henipavirus along with the Hendra virus, which has also caused disease outbreaks.
Pestivirus is a genus of viruses, in the family Flaviviridae. Viruses in the genus Pestivirus infect mammals, including members of the family Bovidae and the family Suidae. There are 11 species in this genus. Diseases associated with this genus include: hemorrhagic syndromes, abortion, and fatal mucosal disease.

A viral envelope is the outermost layer of many types of viruses. It protects the genetic material in their life cycle when traveling between host cells. Not all viruses have envelopes. A viral envelope protein or E protein is a protein in the envelope, which may be acquired by the capsid from an infected host cell.
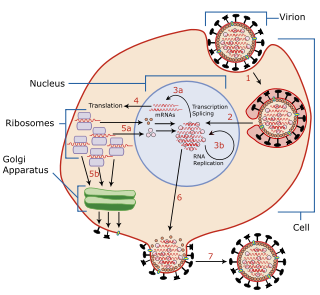
Viral entry is the earliest stage of infection in the viral life cycle, as the virus comes into contact with the host cell and introduces viral material into the cell. The major steps involved in viral entry are shown below. Despite the variation among viruses, there are several shared generalities concerning viral entry.
Pseudotyping is the process of producing viruses or viral vectors in combination with foreign viral envelope proteins. The result is a pseudotyped virus particle, also called a pseudovirus. With this method, the foreign viral envelope proteins can be used to alter host tropism or increase or decrease the stability of the virus particles. Pseudotyped particles do not carry the genetic material to produce additional viral envelope proteins, so the phenotypic changes cannot be passed on to progeny viral particles. In some cases, the inability to produce viral envelope proteins renders the pseudovirus replication incompetent. In this way, the properties of dangerous viruses can be studied in a lower risk setting.
Cedar virus, officially Cedar henipavirus, is a henipavirus known to be harboured by Pteropus spp. Infectious virus was isolated from the urine of a mixed Pteropus alecto and P. poliocephalus in Queensland, Australia in 2009. Unlike the Nipah and Hendra virus, Cedar virus infection does not lead to obvious disease in vivo. Infected animals mounted effective immune responses and seroconverted in challenge studies.

Pneumoviridae is a family of negative-strand RNA viruses in the order Mononegavirales. Humans, cattle, and rodents serve as natural hosts. Respiratory tract infections are associated with member viruses such as human respiratory syncytial virus. There are five species in the family which are divided between the genera Metapneumovirus and Orthopneumovirus. The family used to be considered as a sub-family of Paramyxoviridae, but has been reclassified as of 2016.
Mammalian orthoreovirus (MRV) is a double-stranded RNA virus. It is a part of the family Reoviridae, as well as the subfamily Spinareovirinae. As seen in the name, the Mammalian Ortheoreovirus infects numerous mammalian species and vertebrates which serve as natural hosts. Some diseases that occur as a result of this virus or are associated with this virus include mild upper respiratory illness, and gastrointestinal illness. Examples of these are: upper respiratory tract syndromes, gastroenteritis, biliary atresia, obstructive hydrocephalus, jaundice, alopecia, conjunctivitis, and ‘oily hair’ associated with steatorrhea.

A Nipah virus infection is a viral infection caused by the Nipah virus. Symptoms from infection vary from none to fever, cough, headache, shortness of breath, and confusion. This may worsen into a coma over a day or two, and 50 to 75% of those infected die. Complications can include inflammation of the brain and seizures following recovery.
West Caucasian bat lyssavirus (WCBL) is a member of genus Lyssavirus, family Rhabdoviridae and order Mononegavirales. This virus was first isolated from Miniopterus schreibersii, in the western Caucasus Mountains of southeastern Europe in 2002. WCBL is the most divergent form of Lyssavirus, and is found in Miniopterus bats (insectivorous), Rousettus aegyptiacus, and Eidolon helvum. The latter two are both fruit bats. The virus is fragile as it can be inactivated by UV light and chemicals, such as ether, chloroform, and bleach. WCBL has not been known to infect humans thus far.
Bat mumps orthorubulavirus, formerly Bat mumps rubulavirus (BMV), is a member of genus Orthorubulavirus, family Paramyxoviridae, and order Mononegavirales. Paramyxoviridae viruses were first isolated from bats using heminested PCR with degenerate primers. This process was then followed by Sanger sequencing. A specific location of this virus is not known because it was isolated from bats worldwide. Although multiple paramyxoviridae viruses have been isolated worldwide, BMV specifically has not been isolated thus far. However, BMV was detected in African fruit bats, but no infectious form has been isolated to date. It is known that BMV is transmitted through saliva in the respiratory system of bats. While the virus was considered its own species for a few years, phylogenetic analysis has since shown that it is a member of Mumps orthorubulavirus.
Rio Negro virus is an alphavirus that was first isolated in Argentina in 1980. The virus was first called Ag80-663 but was renamed to Rio Negro virus in 2005. It is a former member of the Venezuelan equine encephalitis complex (VEEC), which are a group of alphaviruses in the Americas that have the potential to emerge and cause disease. Río Negro virus was recently reclassified as a distinct species. Closely related viruses include Mucambo virus and Everglades virus.

Mòjiāng virus(MojV), officially Mojiang henipavirus, is a virus in the family Paramyxoviridae. Based on phylogenetics, Mòjiāng virus is placed in the genus Henipavirus or described as a henipa-like virus. Antibodies raised against Mòjiāng virus glycoproteins are serologically distinct from other henipaviruses (among which higher cross-reactivity is observed).
