Related Research Articles
The genotype is the part of the genetic makeup of a cell, and therefore of any individual, which determines one of its characteristics (phenotype). The term was coined by the Danish botanist, plant physiologist and geneticist Wilhelm Johannsen in 1903.

In genetics, dominance is the phenomenon of one variant (allele) of a gene on a chromosome masking or overriding the effect of a different variant of the same gene on the other copy of the chromosome. The first variant is termed dominant and the second recessive. This state of having two different variants of the same gene on each chromosome is originally caused by a mutation in one of the genes, either new or inherited. The terms autosomal dominant or autosomal recessive are used to describe gene variants on non-sex chromosomes (autosomes) and their associated traits, while those on sex chromosomes (allosomes) are termed X-linked dominant, X-linked recessive or Y-linked; these have an inheritance and presentation pattern that depends on the sex of both the parent and the child. Since there is only one copy of the Y chromosome, Y-linked traits cannot be dominant nor recessive. Additionally, there are other forms of dominance such as incomplete dominance, in which a gene variant has a partial effect compared to when it is present on both chromosomes, and co-dominance, in which different variants on each chromosome both show their associated traits.

Population genetics is a subfield of genetics that deals with genetic differences within and between populations, and is a part of evolutionary biology. Studies in this branch of biology examine such phenomena as adaptation, speciation, and population structure.
Genetic linkage is the tendency of DNA sequences that are close together on a chromosome to be inherited together during the meiosis phase of sexual reproduction. Two genetic markers that are physically near to each other are unlikely to be separated onto different chromatids during chromosomal crossover, and are therefore said to be more linked than markers that are far apart. In other words, the nearer two genes are on a chromosome, the lower the chance of recombination between them, and the more likely they are to be inherited together. Markers on different chromosomes are perfectly unlinked.

In biology, polymorphism is the occurrence of two or more clearly different morphs or forms, also referred to as alternative phenotypes, in the population of a species. To be classified as such, morphs must occupy the same habitat at the same time and belong to a panmictic population.
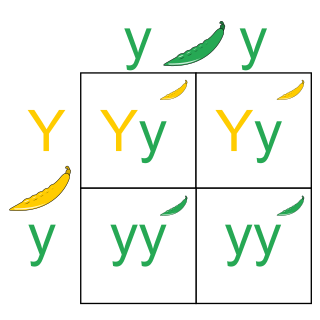
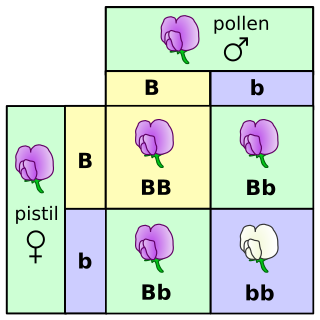
The Punnett square is a square diagram that is used to predict the genotypes of a particular cross or breeding experiment. It is named after Reginald C. Punnett, who devised the approach. The diagram is used by biologists to determine the probability of an offspring having a particular genotype. The Punnett square is a tabular summary of possible combinations of maternal alleles with paternal alleles. These tables can be used to examine the genotypical outcome probabilities of the offspring of a single trait (allele), or when crossing multiple traits from the parents. The Punnett square is a visual representation of Mendelian inheritance. It is important to understand the terms "heterozygous", "homozygous", "double heterozygote", "dominant allele" and "recessive allele" when using the Punnett square method. For multiple traits, using the "forked-line method" is typically much easier than the Punnett square. Phenotypes may be predicted with at least better-than-chance accuracy using a Punnett square, but the phenotype that may appear in the presence of a given genotype can in some instances be influenced by many other factors, as when polygenic inheritance and/or epigenetics are at work.

Cat coat genetics determine the coloration, pattern, length, and texture of feline fur. Understanding how is challenging because many genes are involved. The variations among cat coats are physical properties and should not be confused with cat breeds. A cat may display the coat of a certain breed without actually being that breed. For example, a Siberian could wear point coloration, the stereotypical coat of a Siamese.

Bay is a hair coat color of horses, characterized by a reddish-brown or brown body color with a black point coloration of the mane, tail, ear edges, and lower legs. Bay is one of the most common coat colors in many horse breeds.

Equine coat color genetics determine a horse's coat color. Many colors are possible, but all variations are produced by changes in only a few genes. Extension and agouti are particularly well-known genes with dramatic effects. Differences at the agouti gene determine whether a horse is bay or black, and a change to the extension gene can make a horse chestnut instead. Most domestic horses have a variant of the dun gene which saturates the coat with color so that they are bay, black, or chestnut instead of dun, grullo, or red dun. A mutation called cream is responsible for palomino, buckskin, and cremello horses. Pearl, champagne and silver dapple also lighten the coat, and sometimes the skin and eyes as well. Genes that affect the distribution of melanocytes create patterns of white such as in roan, pinto, leopard, white, and even white markings. Finally, the gray gene causes premature graying, slowly adding white hairs over the course of several years until the horse looks white. Some of these patterns have complex interactions.

Non-Mendelian inheritance is any pattern of inheritance in which traits do not segregate in accordance with Mendel's laws. These laws describe the inheritance of traits linked to single genes on chromosomes in the nucleus. In Mendelian inheritance, each parent contributes one of two possible alleles for a trait. If the genotypes of both parents in a genetic cross are known, Mendel's laws can be used to determine the distribution of phenotypes expected for the population of offspring. There are several situations in which the proportions of phenotypes observed in the progeny do not match the predicted values.
Genetic architecture is the underlying genetic basis of a phenotypic trait and its variational properties. Phenotypic variation for quantitative traits is, at the most basic level, the result of the segregation of alleles at quantitative trait loci (QTL). Environmental factors and other external influences can also play a role in phenotypic variation. Genetic architecture is a broad term that can be described for any given individual based on information regarding gene and allele number, the distribution of allelic and mutational effects, and patterns of pleiotropy, dominance, and epistasis.
A "polygene” or "multiple gene inheritance" is a member of a group of non-epistatic genes that interact additively to influence a phenotypic trait. The term "monozygous" is usually used to refer to a hypothetical gene as it is often difficult to characterise the effect of an individual gene from the effects of other genes and the environment on a particular phenotype. Advances in statistical methodology and high throughput sequencing are, however, allowing researchers to locate candidate genes for the trait. In the case that such a gene is identified, it is referred to as a quantitative trait locus (QTL). These genes are generally pleiotropic as well. The genes that contribute to type 2 diabetes are thought to be mostly polygenes. In July 2016, scientists reported identifying a set of 355 genes from the last universal common ancestor (LUCA) of all organisms living on Earth.
In genetics, transgressive segregation is the formation of extreme phenotypes, or transgressive phenotypes, observed in segregated hybrid populations compared to phenotypes observed in the parental lines. The appearance of these trangressive (extreme) phenotypes can be either positive or negative in terms of fitness. If both parents' favorable alleles come together, it will result in a hybrid having a higher fitness than the two parents. The hybrid species will show more genetic variation and variation in gene expression than their parents. As a result, the hybrid species will have some traits that are transgressive (extreme) in nature. Transgressive segregation can allow a hybrid species to populate different environments/niches in which the parent species do not reside, or compete in the existing environment with the parental species.
Hermann J. Muller (1890–1967), who was a 1946 Nobel Prize winner, coined the terms amorph, hypomorph, hypermorph, antimorph and neomorph to classify mutations based on their behaviour in various genetic situations, as well as gene interaction between themselves. These classifications are still widely used in Drosophila genetics to describe mutations. For a more general description of mutations, see mutation, and for a discussion of allele interactions, see dominance relationship.

The genetic basis of coat colour in the Labrador Retriever has been found to depend on several distinct genes. The interplay among these genes is broadly used as an example of epistasis.

Seal brown is a hair coat color of horses characterized by a near-black body color; with black points, the mane, tail and legs; but also reddish or tan areas around the eyes, muzzle, behind the elbow and in front of the stifle. The term is not to be confused with "brown", which is used by some breed registries to refer to either a seal brown horse or to a dark bay without the additional characteristics of seal brown genetics.
Additive genetic effects occur when two or more genes source a single contribution to the final phenotype, or when alleles of a single gene combine so that their combined effects equal the sum of their individual effects. Non-additive genetic effects involve dominance or epistasis.

Epistasis is a phenomenon in genetics in which the effect of a gene mutation is dependent on the presence or absence of mutations in one or more other genes, respectively termed modifier genes. In other words, the effect of the mutation is dependent on the genetic background in which it appears. Epistatic mutations therefore have different effects on their own than when they occur together. Originally, the term epistasis specifically meant that the effect of a gene variant is masked by that of a different gene.

Modern dog breeds have a wide range of coat colors, patterns, textures and lengths. Knowledge of the genetics of canine coat coloring and patterning and coat texturing and length has improved a great deal in recent years.
The agouti gene (ASIP) is responsible for variations in color in many species. Agouti works with extension to regulate the color of melanin which is produced in hairs. The agouti protein causes red to yellow pheomelanin to be produced, while the competing molecule α-MSH signals production of brown to black eumelanin. In wildtype mice, alternating cycles of agouti and α-MSH production cause agouti coloration. Each hair has bands of yellow which grew during agouti production, and black which grew during α-MSH production. Wildtype mice also have light-colored bellies. The hairs there are a creamy color the whole length because the agouti protein was produced the whole time the hairs were growing.
References
- ↑ Rieger, R.; Michaelis, A.; Green, M.M. (1968), A glossary of genetics and cytogenetics: Classical and molecular, New York: Springer-Verlag, ISBN 9780387076683