
Carbon dioxide is a chemical compound with the chemical formula CO2. It is made up of molecules that each have one carbon atom covalently double bonded to two oxygen atoms. It is found in the gas state at room temperature, and as the source of available carbon in the carbon cycle, atmospheric CO2 is the primary carbon source for life on Earth. In the air, carbon dioxide is transparent to visible light but absorbs infrared radiation, acting as a greenhouse gas. Carbon dioxide is soluble in water and is found in groundwater, lakes, ice caps, and seawater. When carbon dioxide dissolves in water, it forms carbonate and mainly bicarbonate, which causes ocean acidification as atmospheric CO2 levels increase.

Photosynthesis is a biological process used by many cellular organisms to convert light energy into chemical energy, which is stored in organic compounds that can later be metabolized through cellular respiration to fuel the organism's activities. The term usually refers to oxygenic photosynthesis, where oxygen is produced as a byproduct, and some of the chemical energy produced is stored in carbohydrate molecules such as sugars, starch, glycogen and cellulose, which are synthesized from endergonic reaction of carbon dioxide with water. Most plants, algae and cyanobacteria perform photosynthesis; such organisms are called photoautotrophs. Photosynthesis is largely responsible for producing and maintaining the oxygen content of the Earth's atmosphere, and supplies most of the biological energy necessary for complex life on Earth.

In botany, a stoma, also called a stomate, is a pore found in the epidermis of leaves, stems, and other organs, that controls the rate of gas exchange. The pore is bordered by a pair of specialized parenchyma cells known as guard cells that regulate the size of the stomatal opening.

Gas exchange is the physical process by which gases move passively by diffusion across a surface. For example, this surface might be the air/water interface of a water body, the surface of a gas bubble in a liquid, a gas-permeable membrane, or a biological membrane that forms the boundary between an organism and its extracellular environment.

Crassulacean acid metabolism, also known as CAM photosynthesis, is a carbon fixation pathway that evolved in some plants as an adaptation to arid conditions that allows a plant to photosynthesize during the day, but only exchange gases at night. In a plant using full CAM, the stomata in the leaves remain shut during the day to reduce evapotranspiration, but they open at night to collect carbon dioxide and allow it to diffuse into the mesophyll cells. The CO2 is stored as four-carbon malic acid in vacuoles at night, and then in the daytime, the malate is transported to chloroplasts where it is converted back to CO2, which is then used during photosynthesis. The pre-collected CO2 is concentrated around the enzyme RuBisCO, increasing photosynthetic efficiency. This mechanism of acid metabolism was first discovered in plants of the family Crassulaceae.

C4 carbon fixation or the Hatch–Slack pathway is one of three known photosynthetic processes of carbon fixation in plants. It owes the names to the 1960s discovery by Marshall Davidson Hatch and Charles Roger Slack that some plants, when supplied with 14CO2, incorporate the 14C label into four-carbon molecules first.

Photorespiration (also known as the oxidative photosynthetic carbon cycle or C2 cycle) refers to a process in plant metabolism where the enzyme RuBisCO oxygenates RuBP, wasting some of the energy produced by photosynthesis. The desired reaction is the addition of carbon dioxide to RuBP (carboxylation), a key step in the Calvin–Benson cycle, but approximately 25% of reactions by RuBisCO instead add oxygen to RuBP (oxygenation), creating a product that cannot be used within the Calvin–Benson cycle. This process lowers the efficiency of photosynthesis, potentially lowering photosynthetic output by 25% in C3 plants. Photorespiration involves a complex network of enzyme reactions that exchange metabolites between chloroplasts, leaf peroxisomes and mitochondria.
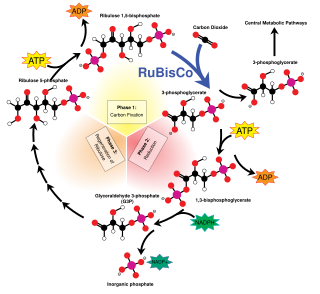
C3 carbon fixation is the most common of three metabolic pathways for carbon fixation in photosynthesis, the other two being C4 and CAM. This process converts carbon dioxide and ribulose bisphosphate (RuBP, a 5-carbon sugar) into two molecules of 3-phosphoglycerate through the following reaction:
The light compensation point (Ic) is the light intensity on the light curve where the rate of photosynthesis exactly matches the rate of cellular respiration. At this point, the uptake of CO2 through photosynthetic pathways is equal to the respiratory release of carbon dioxide, and the uptake of O2 by respiration is equal to the photosynthetic release of oxygen. The concept of compensation points in general may be applied to other photosynthetic variables, the most important being that of CO2 concentration – CO2 compensation point (Γ).Interval of time in day time when light intensity is low due to which net gaseous exchange is zero is called as compensation point.
Moisture stress is a form of abiotic stress that occurs when the moisture of plant tissues is reduced to suboptimal levels. Water stress occurs in response to atmospheric and soil water availability when the transpiration rate exceeds the rate of water uptake by the roots and cells lose turgor pressure. Moisture stress is described by two main metrics, water potential and water content.
Ecophysiology, environmental physiology or physiological ecology is a biological discipline that studies the response of an organism's physiology to environmental conditions. It is closely related to comparative physiology and evolutionary physiology. Ernst Haeckel's coinage bionomy is sometimes employed as a synonym.

Photoinhibition is light-induced reduction in the photosynthetic capacity of a plant, alga, or cyanobacterium. Photosystem II (PSII) is more sensitive to light than the rest of the photosynthetic machinery, and most researchers define the term as light-induced damage to PSII. In living organisms, photoinhibited PSII centres are continuously repaired via degradation and synthesis of the D1 protein of the photosynthetic reaction center of PSII. Photoinhibition is also used in a wider sense, as dynamic photoinhibition, to describe all reactions that decrease the efficiency of photosynthesis when plants are exposed to light.
A xerophyte is a species of plant that has adaptations to survive in an environment with little liquid water. Examples are typically desert regions like the Sahara, and places in the Alps or the Arctic. Popular examples of xerophytes are cacti, pineapple and some Gymnosperm plants.

Transpiration is the process of water movement through a plant and its evaporation from aerial parts, such as leaves, stems and flowers. Water is necessary for plants but only a small amount of water taken up by the roots is used for growth and metabolism. The remaining 97–99.5% is lost by transpiration and guttation. Leaf surfaces are dotted with pores called stomata, and in most plants they are more numerous on the undersides of the foliage. The stomata are bordered by guard cells and their stomatal accessory cells that open and close the pore. Transpiration occurs through the stomatal apertures, and can be thought of as a necessary "cost" associated with the opening of the stomata to allow the diffusion of carbon dioxide gas from the air for photosynthesis. Transpiration also cools plants, changes osmotic pressure of cells, and enables mass flow of mineral nutrients and water from roots to shoots. Two major factors influence the rate of water flow from the soil to the roots: the hydraulic conductivity of the soil and the magnitude of the pressure gradient through the soil. Both of these factors influence the rate of bulk flow of water moving from the roots to the stomatal pores in the leaves via the xylem.
Photosynthetic capacity (Amax) is a measure of the maximum rate at which leaves are able to fix carbon during photosynthesis. It is typically measured as the amount of carbon dioxide that is fixed per metre squared per second, for example as μmol m−2 sec−1.

Plant stress measurement is the quantification of environmental effects on plant health. When plants are subjected to less than ideal growing conditions, they are considered to be under stress. Stress factors can affect growth, survival and crop yields. Plant stress research looks at the response of plants to limitations and excesses of the main abiotic factors, and of other stress factors that are important in particular situations. Plant stress measurement usually focuses on taking measurements from living plants. It can involve visual assessments of plant vitality, however, more recently the focus has moved to the use of instruments and protocols that reveal the response of particular processes within the plant
Stomatal conductance, usually measured in mmol m−2 s−1 by a porometer, estimates the rate of gas exchange and transpiration through the leaf stomata as determined by the degree of stomatal aperture.
Thomas D. Sharkey is a plant biochemist who studies gas exchange between plants and the atmosphere. His research has covered (1) carbon metabolism of photosynthesis from carbon dioxide uptake to carbon export from the Calvin-Benson Cycle, (2) isoprene emission from plants, and (3) abiotic stress tolerance. Four guiding questions are: (1) how leaf photosynthesis affects plant yield, (2) does some carbon fixation follow an oxidative pathway that reduces sugar output but stabilizes photosynthesis, (3) why plants make isoprene, and (4) how plants cope with high temperature.

Photosynthesis converts carbon dioxide to carbohydrates via several metabolic pathways that provide energy to an organism and preferentially react with certain stable isotopes of carbon. The selective enrichment of one stable isotope over another creates distinct isotopic fractionations that can be measured and correlated among oxygenic phototrophs. The degree of carbon isotope fractionation is influenced by several factors, including the metabolism, anatomy, growth rate, and environmental conditions of the organism. Understanding these variations in carbon fractionation across species is useful for biogeochemical studies, including the reconstruction of paleoecology, plant evolution, and the characterization of food chains.
Barley is known to be more environmentally-tolerant than other cereal crops, in terms of soil pH, mineral nutrient availability, and water availability. Because of this, much research is being done on barley plants in order to determine whether or not there is a genetic basis for this environmental hardiness.
