
The entorhinal cortex (EC) is an area of the brain located in the medial temporal lobe and functions as a hub in a widespread network for memory, navigation and the perception of time. The EC is the main interface between the hippocampus and neocortex. The EC-hippocampus system plays an important role in declarative (autobiographical/episodic/semantic) memories and in particular spatial memories including memory formation, memory consolidation, and memory optimization in sleep. The EC is also responsible for the pre-processing (familiarity) of the input signals in the reflex nictitating membrane response of classical trace conditioning; the association of impulses from the eye and the ear occurs in the entorhinal cortex.

Henry Gustav Molaison, known widely as H.M., was an American man who had a bilateral medial temporal lobectomy to surgically resect the anterior two thirds of his hippocampi, parahippocampal cortices, entorhinal cortices, piriform cortices, and amygdalae in an attempt to cure his epilepsy. Although the surgery was partially successful in controlling his epilepsy, a severe side effect was that he became unable to form new memories.

The temporal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The temporal lobe is located beneath the lateral fissure on both cerebral hemispheres of the mammalian brain.
Anterograde amnesia is a loss of the ability to create new memories after the event that caused amnesia, leading to a partial or complete inability to recall the recent past, while long-term memories from before the event remain intact. This is in contrast to retrograde amnesia, where memories created prior to the event are lost while new memories can still be created. Both can occur together in the same patient. To a large degree, anterograde amnesia remains a mysterious ailment because the precise mechanism of storing memories is not yet well understood, although it is known that the regions involved are certain sites in the temporal cortex, especially in the hippocampus and nearby subcortical regions.
Astereognosis is the inability to identify an object by active touch of the hands without other sensory input, such as visual or sensory information. An individual with astereognosis is unable to identify objects by handling them, despite intact elementary tactile, proprioceptive, and thermal sensation. With the absence of vision, an individual with astereognosis is unable to identify what is placed in their hand based on cues such as texture, size, spatial properties, and temperature. As opposed to agnosia, when the object is observed visually, one should be able to successfully identify the object.

In cognitive psychology and neuroscience, spatial memory is a form of memory responsible for the recording of information about one's environment and spatial orientation. For example, a person's spatial memory is required in order to navigate around a familiar city, just as a rat's spatial memory is needed to learn the location of food at the end of a maze. It is often argued that in both humans and animals, spatial memories are summarized as a cognitive map.
The entorhinal cortex (EC) is a major part of the hippocampal formation of the brain, and is reciprocally connected with the hippocampus.
An apical dendrite is a dendrite that emerges from the apex of a pyramidal cell. Apical dendrites are one of two primary categories of dendrites, and they distinguish the pyramidal cells from spiny stellate cells in the cortices. Pyramidal cells are found in the prefrontal cortex, the hippocampus, the entorhinal cortex, the olfactory cortex, and other areas. Dendrite arbors formed by apical dendrites are the means by which synaptic inputs into a cell are integrated. The apical dendrites in these regions contribute significantly to memory, learning, and sensory associations by modulating the excitatory and inhibitory signals received by the pyramidal cells.

The parahippocampal gyrus is a grey matter cortical region of the brain that surrounds the hippocampus and is part of the limbic system. The region plays an important role in memory encoding and retrieval. It has been involved in some cases of hippocampal sclerosis. Asymmetry has been observed in schizophrenia.

Brodmann area 35, together with Brodmann area 36, comprise the perirhinal cortex. They are cytoarchitecturally defined temporal regions of the cerebral cortex.
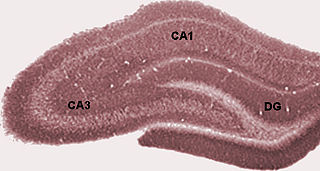
In the brain, the perforant path or perforant pathway provides a connectional route from the entorhinal cortex to all fields of the hippocampal formation, including the dentate gyrus, all CA fields, and the subiculum.

The inferior temporal gyrus is one of three gyri of the temporal lobe and is located below the middle temporal gyrus, connected behind with the inferior occipital gyrus; it also extends around the infero-lateral border on to the inferior surface of the temporal lobe, where it is limited by the inferior sulcus. This region is one of the higher levels of the ventral stream of visual processing, associated with the representation of complex object features, such as global shape. It may also be involved in face perception, and in the recognition of numbers.
The perirhinal cortex is a cortical region in the medial temporal lobe that is made up of Brodmann areas 35 and 36. It receives highly processed sensory information from all sensory regions, and is generally accepted to be an important region for memory. It is bordered caudally by postrhinal cortex or parahippocampal cortex and ventrally and medially by entorhinal cortex.
The neuroanatomy of memory encompasses a wide variety of anatomical structures in the brain.
Recognition memory, a subcategory of declarative memory, is the ability to recognize previously encountered events, objects, or people. When the previously experienced event is reexperienced, this environmental content is matched to stored memory representations, eliciting matching signals. As first established by psychology experiments in the 1970s, recognition memory for pictures is quite remarkable: humans can remember thousands of images at high accuracy after seeing each only once and only for a few seconds.
Visual object recognition refers to the ability to identify the objects in view based on visual input. One important signature of visual object recognition is "object invariance", or the ability to identify objects across changes in the detailed context in which objects are viewed, including changes in illumination, object pose, and background context.
The study of memory incorporates research methodologies from neuropsychology, human development and animal testing using a wide range of species. The complex phenomenon of memory is explored by combining evidence from many areas of research. New technologies, experimental methods and animal experimentation have led to an increased understanding of the workings of memory.
Distributed practice is a learning strategy, where practice is broken up into a number of short sessions – over a longer period of time. Humans and animals learn items in a list more effectively when they are studied in several sessions spread out over a long period of time, rather than studied repeatedly in a short period of time, a phenomenon called the spacing effect. The opposite, massed practice, consists of fewer, longer training sessions. It is generally a less effective method of learning. For example, when studying for an exam dispersing your studying more frequently over a larger period of time will result in more effective learning than intense study the night before.
In psychology, associative memory is defined as the ability to learn and remember the relationship between unrelated items. This would include, for example, remembering the name of someone or the aroma of a particular perfume. This type of memory deals specifically with the relationship between these different objects or concepts. A normal associative memory task involves testing participants on their recall of pairs of unrelated items, such as face-name pairs. Associative memory is a declarative memory structure and episodically based.
Elizabeth A. Buffalo is Professor of Physiology and Biophysics at the University of Washington School of Medicine and Chief of the Neuroscience Division at the Washington National Primate Research Center. She is known for her research in the field of neurophysiology pertaining to the role of the hippocampus and medial temporal lobe structures in learning and memory and in spatial representation and navigation.

{kind=link}