Related Research Articles

Cystic fibrosis transmembrane conductance regulator (CFTR) is a membrane protein and anion channel in vertebrates that is encoded by the CFTR gene.

Chloride channels are a superfamily of poorly understood ion channels specific for chloride. These channels may conduct many different ions, but are named for chloride because its concentration in vivo is much higher than other anions. Several families of voltage-gated channels and ligand-gated channels have been characterized in humans.

Prestin is a protein that is critical to sensitive hearing in mammals. It is encoded by the SLC26A5 gene.
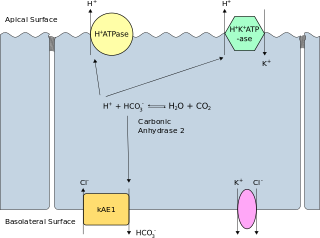
Band 3 anion transport protein, also known as anion exchanger 1 (AE1) or band 3 or solute carrier family 4 member 1 (SLC4A1), is a protein that is encoded by the SLC4A1 gene in humans.
The Transporter Classification Database is an International Union of Biochemistry and Molecular Biology (IUBMB)-approved classification system for membrane transport proteins, including ion channels.

The epithelial sodium channel(ENaC), (also known as amiloride-sensitive sodium channel) is a membrane-bound ion channel that is selectively permeable to sodium ions (Na+). It is assembled as a heterotrimer composed of three homologous subunits α or δ, β, and γ, These subunits are encoded by four genes: SCNN1A, SCNN1B, SCNN1G, and SCNN1D. The ENaC is involved primarily in the reabsorption of sodium ions at the collecting ducts of the kidney's nephrons. In addition to being implicated in diseases where fluid balance across epithelial membranes is perturbed, including pulmonary edema, cystic fibrosis, COPD and COVID-19, proteolyzed forms of ENaC function as the human salt taste receptor.
Pendrin is an anion exchange protein that in humans is encoded by the SLC26A4 gene . Pendrin was initially identified as a sodium-independent chloride-iodide exchanger with subsequent studies showing that it also accepts formate and bicarbonate as substrates. Pendrin is similar to the Band 3 transport protein found in red blood cells. Pendrin is the protein which is mutated in Pendred syndrome, which is an autosomal recessive disorder characterized by sensorineural hearing loss, goiter and a partial organification problem detectable by a positive perchlorate test.
Translocase is a general term for a protein that assists in moving another molecule, usually across a cell membrane. These enzymes catalyze the movement of ions or molecules across membranes or their separation within membranes. The reaction is designated as a transfer from “side 1” to “side 2” because the designations “in” and “out”, which had previously been used, can be ambiguous. Translocases are the most common secretion system in Gram positive bacteria.

The sulfate transporter is a solute carrier family protein that in humans is encoded by the SLC26A2 gene. SLC26A2 is also called the diastrophic dysplasia sulfate transporter (DTDST), and was first described by Hästbacka et al. in 1994. A defect in sulfate activation described by Superti-Furga in achondrogenesis type 1B was subsequently also found to be caused by genetic variants in the sulfate transporter gene. This sulfate (SO42−) transporter also accepts chloride, hydroxyl ions (OH−), and oxalate as substrates. SLC26A2 is expressed at high levels in developing and mature cartilage, as well as being expressed in lung, placenta, colon, kidney, pancreas and testis.
Chloride anion exchanger, also known as down-regulated in adenoma, is a protein that in humans is encoded by the SLC26A3 gene.

Anion exchange protein 3 is a membrane transport protein that in humans is encoded by the SLC4A3 gene. AE3 is functionally similar to the Band 3 Cl−/HCO3− exchange protein but it is expressed primarily in brain neurons and in the heart. Like AE2 its activity is sensitive to pH. AE3 mutations have been linked to seizures.

Solute carrier family 26 member 6 is a protein that in humans is encoded by the SLC26A6 gene. It is an anion-exchanger expressed in the apical membrane of the kidney proximal tubule, the apical membranes of the duct cells in the pancreas, and the villi of the duodenum.

Anion exchange transporter is a protein that in humans is encoded by the SLC26A7 gene.
Proteins currently known to belong to the Ni2+-Co2+ Transporter (NiCoT) family (TC# 2.A.52) can be found in organisms ranging from Gram-negative and Gram-positive bacteria to archaea and some eukaryotes. Members of this family catalyze uptake of Ni2+ and/or Co2+ in a proton motive force-dependent process.
Members of the organic solute transporter (OST) family have been characterized from a small bottom feeding species of fish called the little skate, Raja erinacea. Members have also been characterized from humans and mice. The OST family is a member of the larger group of secondary carriers, the APC superfamily.
The amino acid-polyamine-organocation (APC) superfamily is the second largest superfamily of secondary carrier proteins currently known, and it contains several Solute carriers. Originally, the APC superfamily consisted of subfamilies under the transporter classification number. This superfamily has since been expanded to include eighteen different families.
The anion exchanger family is a member of the large APC superfamily of secondary carriers. Members of the AE family are generally responsible for the transport of anions across cellular barriers, although their functions may vary. All of them exchange bicarbonate. Characterized protein members of the AE family are found in plants, animals, insects and yeast. Uncharacterized AE homologues may be present in bacteria. Animal AE proteins consist of homodimeric complexes of integral membrane proteins that vary in size from about 900 amino acyl residues to about 1250 residues. Their N-terminal hydrophilic domains may interact with cytoskeletal proteins and therefore play a cell structural role. Some of the currently characterized members of the AE family can be found in the Transporter Classification Database.
The potassium (K+) uptake permease (KUP) family (TC# 2.A.72) is a member of the APC superfamily of secondary carriers. Proteins of the KUP/HAK/KT family include the KUP (TrkD) protein of E. coli and homologues in both Gram-positive and Gram-negative bacteria. High affinity (20 μM) K+ uptake systems (Hak1, TC# 2.A.72.2.1) of the yeast Debaryomyces occidentalis as well as the fungus, Neurospora crassa, and several homologues in plants have been characterized. Arabidopsis thaliana and other plants possess multiple KUP family paralogues. While many plant proteins cluster tightly together, the Hak1 proteins from yeast as well as the two Gram-positive and Gram-negative bacterial proteins are distantly related on the phylogenetic tree for the KUP family. All currently classified members of the KUP family can be found in the Transporter Classification Database.
Arsenite resistance (Ars) efflux pumps of bacteria may consist of two proteins, ArsB and ArsA, or of one protein. ArsA proteins have two ATP binding domains and probably arose by a tandem gene duplication event. ArsB proteins all possess twelve transmembrane spanners and may also have arisen by a tandem gene duplication event. Structurally, the Ars pumps resemble ABC-type efflux pumps, but there is no significant sequence similarity between the Ars and ABC pumps. When only ArsB is present, the system operates by a pmf-dependent mechanism, and consequently belongs in TC subclass 2.A. When ArsA is also present, ATP hydrolysis drives efflux, and consequently the system belongs in TC subclass 3.A. ArsB therefore appears twice in the TC system but ArsA appears only once. These pumps actively expel both arsenite and antimonite.
The inorganic phosphate transporter (PiT) family is a group of carrier proteins derived from Gram-negative and Gram-positive bacteria, archaea, and eukaryotes.
References
- ↑ Wong FH, Chen JS, Reddy V, Day JL, Shlykov MA, Wakabayashi ST, Saier MH (2012-01-01). "The amino acid-polyamine-organocation superfamily". Journal of Molecular Microbiology and Biotechnology. 22 (2): 105–13. doi: 10.1159/000338542 . PMID 22627175.
- ↑ Jiang Z, Grichtchenko II, Boron WF, Aronson PS (September 2002). "Specificity of anion exchange mediated by mouse Slc26a6". The Journal of Biological Chemistry. 277 (37): 33963–7. doi: 10.1074/jbc.M202660200 . PMID 12119287.
- ↑ Maeda S, Sugita C, Sugita M, Omata T (March 2006). "Latent nitrate transport activity of a novel sulfate permease-like protein of the cyanobacterium Synechococcus elongatus". The Journal of Biological Chemistry. 281 (9): 5869–76. doi: 10.1074/jbc.M513196200 . PMID 16407232.
- 1 2 3 Ohana E, Shcheynikov N, Yang D, So I, Muallem S (February 2011). "Determinants of coupled transport and uncoupled current by the electrogenic SLC26 transporters". The Journal of General Physiology. 137 (2): 239–51. doi:10.1085/jgp.201010531. PMC 3032377 . PMID 21282402.
- 1 2 Detro-Dassen S, Schänzler M, Lauks H, Martin I, zu Berstenhorst SM, Nothmann D, Torres-Salazar D, Hidalgo P, Schmalzing G, Fahlke C (February 2008). "Conserved dimeric subunit stoichiometry of SLC26 multifunctional anion exchangers". The Journal of Biological Chemistry. 283 (7): 4177–88. doi: 10.1074/jbc.M704924200 . PMID 18073211.
- 1 2 Saier, MH Jr. "2.A.53 The Sulfate Permease (SulP) Family". Transporter Classification Database. Saier Lab Bioinformatics Group / SDSC.
- ↑ Price GD, Woodger FJ, Badger MR, Howitt SM, Tucker L (December 2004). "Identification of a SulP-type bicarbonate transporter in marine cyanobacteria". Proceedings of the National Academy of Sciences of the United States of America. 101 (52): 18228–33. Bibcode:2004PNAS..10118228P. doi: 10.1073/pnas.0405211101 . PMC 539743 . PMID 15596724.
- 1 2 Felce J, Saier MH (2004-01-01). "Carbonic anhydrases fused to anion transporters of the SulP family: evidence for a novel type of bicarbonate transporter". Journal of Molecular Microbiology and Biotechnology. 8 (3): 169–76. doi:10.1159/000085789. PMID 16088218. S2CID 9550955.
- ↑ Schweinfest CW, Spyropoulos DD, Henderson KW, Kim JH, Chapman JM, Barone S, Worrell RT, Wang Z, Soleimani M (December 2006). "slc26a3 (dra)-deficient mice display chloride-losing diarrhea, enhanced colonic proliferation, and distinct up-regulation of ion transporters in the colon". The Journal of Biological Chemistry. 281 (49): 37962–71. doi: 10.1074/jbc.M607527200 . PMID 17001077.
- ↑ Tomatsu H, Takano J, Takahashi H, Watanabe-Takahashi A, Shibagaki N, Fujiwara T (November 2007). "An Arabidopsis thaliana high-affinity molybdate transporter required for efficient uptake of molybdate from soil". Proceedings of the National Academy of Sciences of the United States of America. 104 (47): 18807–12. Bibcode:2007PNAS..10418807T. doi: 10.1073/pnas.0706373104 . PMC 2141858 . PMID 18003916.
- ↑ Ousingsawat J, Schreiber R, Kunzelmann K (June 2012). "Differential contribution of SLC26A9 to Cl(-) conductance in polarized and non-polarized epithelial cells". Journal of Cellular Physiology. 227 (6): 2323–9. doi:10.1002/jcp.22967. PMID 21809345. S2CID 35893617.
As of 2 February 2016, this article is derived in whole or in part from Transporter Classification Database . The copyright holder has licensed the content in a manner that permits reuse under CC BY-SA 3.0 and GFDL. All relevant terms must be followed.The original text was at "2.A.53 The Sulfate Permease (SulP) Family"