Related Research Articles
A restriction enzyme, restriction endonuclease, REase, ENase orrestrictase is an enzyme that cleaves DNA into fragments at or near specific recognition sites within molecules known as restriction sites. Restriction enzymes are one class of the broader endonuclease group of enzymes. Restriction enzymes are commonly classified into five types, which differ in their structure and whether they cut their DNA substrate at their recognition site, or if the recognition and cleavage sites are separate from one another. To cut DNA, all restriction enzymes make two incisions, once through each sugar-phosphate backbone of the DNA double helix.
Gene knockouts are a widely used genetic engineering technique that involves the targeted removal or inactivation of a specific gene within an organism's genome. This can be done through a variety of methods, including homologous recombination, CRISPR-Cas9, and TALENs.
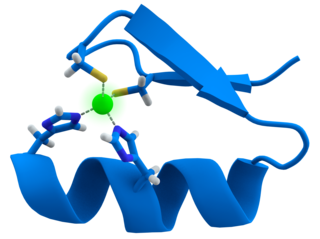
A zinc finger is a small protein structural motif that is characterized by the coordination of one or more zinc ions (Zn2+) which stabilizes the fold. It was originally coined to describe the finger-like appearance of a hypothesized structure from the African clawed frog (Xenopus laevis) transcription factor IIIA. However, it has been found to encompass a wide variety of differing protein structures in eukaryotic cells. Xenopus laevis TFIIIA was originally demonstrated to contain zinc and require the metal for function in 1983, the first such reported zinc requirement for a gene regulatory protein followed soon thereafter by the Krüppel factor in Drosophila. It often appears as a metal-binding domain in multi-domain proteins.
The restriction modification system is found in bacteria and other prokaryotic organisms, and provides a defense against foreign DNA, such as that borne by bacteriophages.

A germline mutation, or germinal mutation, is any detectable variation within germ cells. Mutations in these cells are the only mutations that can be passed on to offspring, when either a mutated sperm or oocyte come together to form a zygote. After this fertilization event occurs, germ cells divide rapidly to produce all of the cells in the body, causing this mutation to be present in every somatic and germline cell in the offspring; this is also known as a constitutional mutation. Germline mutation is distinct from somatic mutation.

C-C chemokine receptor type 5, also known as CCR5 or CD195, is a protein on the surface of white blood cells that is involved in the immune system as it acts as a receptor for chemokines.

A designer baby is a baby whose genetic makeup has been selected or altered, often to exclude a particular gene or to remove genes associated with disease. This process usually involves analysing a wide range of human embryos to identify genes associated with particular diseases and characteristics, and selecting embryos that have the desired genetic makeup; a process known as preimplantation genetic diagnosis. Screening for single genes is commonly practiced, and polygenic screening is offered by a few companies. Other methods by which a baby's genetic information can be altered involve directly editing the genome before birth, which is not routinely performed and only one instance of this is known to have occurred as of 2019, where Chinese twins Lulu and Nana were edited as embryos, causing widespread criticism.

C-X-C chemokine receptor type 4 (CXCR-4) also known as fusin or CD184 is a protein that in humans is encoded by the CXCR4 gene. The protein is a CXC chemokine receptor.
Entry inhibitors, also known as fusion inhibitors, are a class of antiviral drugs that prevent a virus from entering a cell, for example, by blocking a receptor. Entry inhibitors are used to treat conditions such as HIV and hepatitis D.
Zinc-finger nucleases (ZFNs) are artificial restriction enzymes generated by fusing a zinc finger DNA-binding domain to a DNA-cleavage domain. Zinc finger domains can be engineered to target specific desired DNA sequences and this enables zinc-finger nucleases to target unique sequences within complex genomes. By taking advantage of endogenous DNA repair machinery, these reagents can be used to precisely alter the genomes of higher organisms. Alongside CRISPR/Cas9 and TALEN, ZFN is a prominent tool in the field of genome editing.

Gene targeting is a biotechnological tool used to change the DNA sequence of an organism. It is based on the natural DNA-repair mechanism of Homology Directed Repair (HDR), including Homologous Recombination. Gene targeting can be used to make a range of sizes of DNA edits, from larger DNA edits such as inserting entire new genes into an organism, through to much smaller changes to the existing DNA such as a single base-pair change. Gene targeting relies on the presence of a repair template to introduce the user-defined edits to the DNA. The user will design the repair template to contain the desired edit, flanked by DNA sequence corresponding (homologous) to the region of DNA that the user wants to edit; hence the edit is targeted to a particular genomic region. In this way Gene Targeting is distinct from natural homology-directed repair, during which the ‘natural’ DNA repair template of the sister chromatid is used to repair broken DNA. The alteration of DNA sequence in an organism can be useful in both a research context – for example to understand the biological role of a gene – and in biotechnology, for example to alter the traits of an organism.
Zinc finger protein chimera are chimeric proteins composed of a DNA-binding zinc finger protein domain and another domain through which the protein exerts its effect. The effector domain may be a transcriptional activator (A) or repressor (R), a methylation domain (M) or a nuclease (N).
Recombinant adeno-associated virus (rAAV) based genome engineering is a genome editing platform centered on the use of recombinant AAV vectors that enables insertion, deletion or substitution of DNA sequences into the genomes of live mammalian cells. The technique builds on Mario Capecchi and Oliver Smithies' Nobel Prize–winning discovery that homologous recombination (HR), a natural hi-fidelity DNA repair mechanism, can be harnessed to perform precise genome alterations in mice. rAAV mediated genome-editing improves the efficiency of this technique to permit genome engineering in any pre-established and differentiated human cell line, which, in contrast to mouse ES cells, have low rates of HR.
Transcription activator-like effector nucleases (TALEN) are restriction enzymes that can be engineered to cut specific sequences of DNA. They are made by fusing a TAL effector DNA-binding domain to a DNA cleavage domain. Transcription activator-like effectors (TALEs) can be engineered to bind to practically any desired DNA sequence, so when combined with a nuclease, DNA can be cut at specific locations. The restriction enzymes can be introduced into cells, for use in gene editing or for genome editing in situ, a technique known as genome editing with engineered nucleases. Alongside zinc finger nucleases and CRISPR/Cas9, TALEN is a prominent tool in the field of genome editing.

Genome editing, or genome engineering, or gene editing, is a type of genetic engineering in which DNA is inserted, deleted, modified or replaced in the genome of a living organism. Unlike early genetic engineering techniques that randomly inserts genetic material into a host genome, genome editing targets the insertions to site-specific locations. The basic mechanism involved in genetic manipulations through programmable nucleases is the recognition of target genomic loci and binding of effector DNA-binding domain (DBD), double-strand breaks (DSBs) in target DNA by the restriction endonucleases, and the repair of DSBs through homology-directed recombination (HDR) or non-homologous end joining (NHEJ).
Genetic engineering techniques allow the modification of animal and plant genomes. Techniques have been devised to insert, delete, and modify DNA at multiple levels, ranging from a specific base pair in a specific gene to entire genes. There are a number of steps that are followed before a genetically modified organism (GMO) is created. Genetic engineers must first choose what gene they wish to insert, modify, or delete. The gene must then be isolated and incorporated, along with other genetic elements, into a suitable vector. This vector is then used to insert the gene into the host genome, creating a transgenic or edited organism.
HIV/AIDS research includes all medical research that attempts to prevent, treat, or cure HIV/AIDS, as well as fundamental research about the nature of HIV as an infectious agent and AIDS as the disease caused by HIV.
A small proportion of humans show partial or apparently complete innate resistance to HIV, the virus that causes AIDS. The main mechanism is a mutation of the gene encoding CCR5, which acts as a co-receptor for HIV. It is estimated that the proportion of people with some form of resistance to HIV is under 10%.
Off-target genome editing refers to nonspecific and unintended genetic modifications that can arise through the use of engineered nuclease technologies such as: clustered, regularly interspaced, short palindromic repeats (CRISPR)-Cas9, transcription activator-like effector nucleases (TALEN), meganucleases, and zinc finger nucleases (ZFN). These tools use different mechanisms to bind a predetermined sequence of DNA (“target”), which they cleave, creating a double-stranded chromosomal break (DSB) that summons the cell's DNA repair mechanisms and leads to site-specific modifications. If these complexes do not bind at the target, often a result of homologous sequences and/or mismatch tolerance, they will cleave off-target DSB and cause non-specific genetic modifications. Specifically, off-target effects consist of unintended point mutations, deletions, insertions inversions, and translocations.
Paula Cannon is a British geneticist and virologist, Distinguished Professor of Molecular Microbiology & Immunology at the University of Southern California. She is a specialist in gene therapy, hematopoietic stem cells, and human immunodeficiency virus (HIV) with particular interest in gene editing and humanized mice.
References
- ↑ Deeks, S. G.; McCune, J. M. (2010). "Can HIV be cured with stem cell therapy?". Nature Biotechnology. 28 (8): 807–810. doi:10.1038/nbt0810-807. PMID 20697404. S2CID 20312608.
- ↑ Alkhatib, G (2009). "The biology of CCR5 and CXCR4". Current Opinion in HIV and AIDS. 4 (2): 96–103. doi:10.1097/coh.0b013e328324bbec. PMC 2718543 . PMID 19339947.
- 1 2 Hütter, G.; Nowak, D.; Mossner, M.; Ganepola, S.; Müßig, A.; Allers, K.; Thiel, E. (2009). "Long-term control of HIV by CCR5 Delta32/Delta32 stem-cell transplantation". New England Journal of Medicine. 360 (7): 692–698. doi: 10.1056/nejmoa0802905 . PMID 19213682. S2CID 14905671.
- ↑ Carroll, D (2008). "Progress and prospects: zinc-finger nucleases as gene therapy agents". Gene Therapy. 15 (22): 1463–1468. doi:10.1038/gt.2008.145. PMC 2747807 . PMID 18784746.
- ↑ Perez, E. E.; Wang, J.; Miller, J. C.; Jouvenot, Y.; Kim, K. A.; Liu, O.; June, C.H. (2008). "Establishment of HIV-1 resistance in CD4+ T cells by genome editing using zinc-finger nucleases". Nature Biotechnology. 26 (7): 808–816. doi:10.1038/nbt1410. PMC 3422503 . PMID 18587387.
- ↑ Chung, J.; Rossi, J. J.; Jung, U. (2011). "Current progress and challenges in HIV gene therapy". Future Virology. 6 (11): 1319–1328. doi:10.2217/fvl.11.113. PMC 3383045 . PMID 22754586.
- 1 2 Lai, Y. CCR5-targeted hematopoietic stem cell gene approaches for HIV disease: Current progress and future prospects Current Stem Cell Research and Therapy, 2012; 7 (4), pp. 310-317.
- ↑ De Silva, E., Stumpf, Michael P.H. (2004). "HIV and the CCR5-D32 Resistance Allele". FEMS Microbiology Letters. 241 (1): 1–12. doi: 10.1016/j.femsle.2004.09.040 . PMID 15556703.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - 1 2 3 4 5 6 7 8 Carroll, D (2011). "Genome engineering with zinc-finger nucleases". Genetics. 188 (4): 773–782. doi:10.1534/genetics.111.131433. PMC 3176093 . PMID 21828278.
- ↑ Durand, Christine. M, Siliciano, Robert F. (2014). "Dual Zinc-Finger Nucleases Block HIV Infection". Blood. 123 (1): 636–646. doi: 10.1182/blood-2013-11-535542 . PMID 24385492.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - 1 2 3 4 5 Urnov, F. D.; Rebar, E. J.; Holmes, M. C.; Zhang, H. S.; Gregory, P. D. (2010). "Genome editing with engineered zinc finger nucleases". Nature Reviews Genetics. 11 (9): 636–646. doi:10.1038/nrg2842. PMID 20717154. S2CID 205484701.
- 1 2 Urnov, F. D.; Miller, J. C.; Lee; Beausejour; Rock, J. M.; Augustus, S.; Holmes, M. C. (2005). "Highly efficient endogenous human gene correction using designed zinc-finger nucleases". Nature. 435 (7042): 646–651. Bibcode:2005Natur.435..646U. doi:10.1038/nature03556. PMID 15806097. S2CID 4390010.
- ↑ Wayengera, M. "Proviral HIV-genome-wide and pol-gene specific zinc finger nucleases: usability for targeted HIV gene therapy. Theor Biol Med Model, 2011; 8, pp26.
- ↑ Wayengera, M.Zinc finger arrays binding human papillomavirus types 16 and 18 genomic DNA: precursors of gene-therapeutics for in-situ reversal of associated cervical neoplasia.Theor Biol Med Model, (2011), 9, pp30.
- ↑ Wayengera, M.Identity of zinc finger nucleases with specificity to herpes simplex virus type II genomic DNA: novel HSV-2 vaccine/therapy precursors.Theor Biol Med Model, (2011), 8, pp23.
- ↑ Wayengera, M (2003). "HIV and Gene Therapy: The proposed [R-M enzymatic] model for a gene therapy against HIV". Makerere Med J. 38: 28–30.
- ↑ Wayengera, M; Kajumbula, H; Byarugaba, W (2007). "Frequency and site mapping of HIV-1/SIVcpz, HIV-2/SIVsmm and Other SIV gene sequence cleavage by various bacteria restriction enzymes: Precursors for a novel HIV inhibitory product". Afr J Biotechnol. 6 (10): 1225–1232.
- ↑ Wayengera M, Kajumbula H, Byarugaba W: Identification of restriction endonuclease with potential ability to cleave the HSV-2 genome: inherent potential for biosynthetic versus live microbicides. Theor Biol Med Model. 2008, 5:18.
- ↑ Wayengera, M (2008). "Pre-Integration gene slicing (PRINT-GSX) as an alternate or complementary gene therapy modem to RNA interference". J Appl Biol Sci. 1 (2): 56–63.
- ↑ Wayengera M: Diverting primary HIV entry and replication to vaginal commensal lactobacillus expressing R-M nucleic enzymatic peptides with potent activity at cleaving proviral DNA as a novel HIV live microbicide strategy. Microbicide- New Delhi, India 2008. Abs-10.
- ↑ Wayengera M: Preparing for a Phase 1 Preclinical trial of VRX-SMR: a Lentiviral Vector transduced with restriction enzymes cleaving HIV proviral DNA as a therapeutic vaccine: Opportunities and Challenges. Vaccine Congress -Amsterdam, Netherlands 2007, :24OR.
- ↑ Wayengera M: xREPLAB: A recombinant lactobacillus strain producing restriction enzymes with potent activity against HIV proviral DNA as a Live Microbicide Strategy. AIDS vaccine- Washington, Seattle 2007, :P05-01.
- ↑ Wayengera, M (2007). "PREX-1979: Modeling the first ever prototype of could be a 5th generation of Microbicides for preventing HIV infection among high-risk women". Afr J Biotechnol. 6 (10): 1221–1224.
- 1 2 Urnov, F. D., Rebar, E. J., Holmes, M. C., Zhang, H. S., & Gregory, P. D. (2010). "Genome Editing with Engineered Zinc Finger Nucleases". Nature Reviews Genetics. 11 (9): 636–646. doi:10.1038/nrg2842. PMID 20717154. S2CID 205484701.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - 1 2 3 4 5 6 7 8 9 10 11 Pavletich, N. P.; Pabo, C. O. (1991). "Zinc finger-DNA recognition: crystal structure of a Zif268-DNA complex at 2.1 A.". Science. 252 (5007): 809–817. Bibcode:1991Sci...252..809P. doi:10.1126/science.2028256. PMID 2028256. S2CID 38000717.
- 1 2 3 4 5 6 7 8 9 10 11 Klug, A (2005). "The discovery of zinc fingers and their development for practical applications in gene regulation". Proceedings of the Japan Academy, Series B. 81 (4): 87–102. Bibcode:2005PJAB...81...87K. doi: 10.2183/pjab.81.87 .
- ↑ Frankel, A. D.; Berg, J. M.; Pabo, C. O. (1987). "Metal-dependent folding of a single zinc finger from transcription factor IIIA". Proceedings of the National Academy of Sciences. 84 (14): 4841–4845. Bibcode:1987PNAS...84.4841F. doi: 10.1073/pnas.84.14.4841 . PMC 305201 . PMID 3474629.
- 1 2 Lee, M. S.; Gippert, G. P.; Soman, K. V.; Case, D. A.; Wright, P. E. (1989). "Three-dimensional solution structure of a single zinc finger DNA-binding domain". Science. 245 (4918): 635–637. Bibcode:1989Sci...245..635L. doi:10.1126/science.2503871. PMID 2503871.
- 1 2 3 4 Klug, A.; Schwabe, J. W. (1995). "Protein motifs 5. Zinc fingers". The FASEB Journal. 9 (8): 597–604. doi: 10.1096/fasebj.9.8.7768350 . PMID 7768350. S2CID 24135293.
- 1 2 3 4 Stone, D.; Kiem, H. P.; Jerome, K. R. (2013). "Targeted gene disruption to cure HIV". Curr Opin HIV AIDS. 8 (3): 217–23. doi:10.1097/COH.0b013e32835f736c. PMC 4226633 . PMID 23478911.
- 1 2 Coakley, E.; Petropoulos, C.J.; Whitcomb, J.M. (2005). "Assessing ch vbgemokine co-receptor usage in HIV". Curr. Opin. Infect. Dis. 18 (1): 9–15. doi:10.1097/00001432-200502000-00003. PMID 15647694. S2CID 30923492.
- ↑ Wilen, C.B.; Wang, J.; Tilton, J.C.; et al. (2011). "Engineering HIV-resistant humant CD4+ T cells with CXCR4-specific zinc-finger nucleases". PLOS Pathogens. 7 (4): e1002020. doi: 10.1371/journal.ppat.1002020 . PMC 3077364 . PMID 21533216.
- ↑ Didigu, C.A.; Wilen, C.B.; Wang, J. (2013). "Simultaneous zinc-finger nuclease editing of the HIV coreceptors ccr5 and cxcr4 protects CD4+ T cells from HIV-1 infection". Blood. 123 (1): 61–69. doi:10.1182/blood-2013-08-521229. PMC 3879906 . PMID 24162716.
- 1 2 Barton, K. M.; Burch, B. D.; Soriano-Sarabia, N.; Margolis, D. M. (2013). "Prospects for treatment of latent HIV". Clinical Pharmacology and Therapeutics. 93 (1): 46–56. doi:10.1038/clpt.2012.202. PMC 3942883 . PMID 23212106.
- 1 2 3 4 5 6 Cathomen, T., & Joung, J. K.. Zinc-finger nucleases: the next generation emerges. Molecular Therapy, (2008) 16(7), pp 1200-1207.
- 1 2 Levine, B. L.; Humeau, L. M.; Boyer, J.; MacGregor, R. R.; Rebello, T.; Lu, X.; June, C. H. (2006). "Gene transfer in humans using a conditionally replicating lentiviral vector". Proceedings of the National Academy of Sciences. 103 (46): 17372–17377. Bibcode:2006PNAS..10317372L. doi: 10.1073/pnas.0608138103 . PMC 1635018 . PMID 17090675.
- ↑ Varela-Rohena, A.; Carpenito, C.; Perez, E. E.; Richardson, M.; Parry, R. V.; Milone, M.; Riley, J. L. (2008). "Genetic engineering of T cells for adoptive immunotherapy". Immunologic Research. 42 (1–3): 166–181. doi:10.1007/s12026-008-8057-6. PMC 2699549 . PMID 18841331.
- 1 2 3 4 5 Rosenberg, T. "The man who had HIV and now does not". New York Magazine. Retrieved January 2013.
- ↑ Hütter G, Ganepola S (2011). "Eradication of HIV by transplantation of CCR5-deficient hematopoietic stem cells". The Scientific World Journal . 11: 1068–1076. doi: 10.1100/tsw.2011.102 . PMC 5720062 . PMID 21552772.