
The central nervous system (CNS) is the part of the nervous system consisting primarily of the brain and spinal cord. The CNS is so named because the brain integrates the received information and coordinates and influences the activity of all parts of the bodies of bilaterally symmetric and triploblastic animals—that is, all multicellular animals except sponges and diploblasts. It is a structure composed of nervous tissue positioned along the rostral to caudal axis of the body and may have an enlarged section at the rostral end which is a brain. Only arthropods, cephalopods and vertebrates have a true brain, though precursor structures exist in onychophorans, gastropods and lancelets.

The thalamus is a large mass of gray matter located in the dorsal part of the diencephalon. Nerve fibers project out of the thalamus to the cerebral cortex in all directions, allowing hub-like exchanges of information. It has several functions, such as the relaying of sensory signals, including motor signals to the cerebral cortex and the regulation of consciousness, sleep, and alertness.
The development of the nervous system, or neural development (neurodevelopment), refers to the processes that generate, shape, and reshape the nervous system of animals, from the earliest stages of embryonic development to adulthood. The field of neural development draws on both neuroscience and developmental biology to describe and provide insight into the cellular and molecular mechanisms by which complex nervous systems develop, from nematodes and fruit flies to mammals.

In the developing chordate, the neural tube is the embryonic precursor to the central nervous system, which is made up of the brain and spinal cord. The neural groove gradually deepens as the neural fold become elevated, and ultimately the folds meet and coalesce in the middle line and convert the groove into the closed neural tube. In humans, neural tube closure usually occurs by the fourth week of pregnancy.

Sonic hedgehog protein(SHH) is encoded for by the SHH gene. The protein is named after the character Sonic the Hedgehog.

In the human brain, the diencephalon is a division of the forebrain. It is situated between the telencephalon and the midbrain. The diencephalon has also been known as the 'tweenbrain in older literature. It consists of structures that are on either side of the third ventricle, including the thalamus, the hypothalamus, the epithalamus and the subthalamus.

Neurulation refers to the folding process in vertebrate embryos, which includes the transformation of the neural plate into the neural tube. The embryo at this stage is termed the neurula.
Neuromeres are morphologically or molecularly defined transient segments of the early developing brain. Rhombomeres are such segments that make up the rhombencephalon or hindbrain. More controversially, some argue that there exist early developmental segments that give rise to structures of the midbrain (mesomeres) and forebrain (prosomeres).

The metencephalon is the embryonic part of the hindbrain that differentiates into the pons and the cerebellum. It contains a portion of the fourth ventricle and the trigeminal nerve, abducens nerve, facial nerve, and a portion of the vestibulocochlear nerve.
The primitive node is the organizer for gastrulation in most amniote embryos. In birds it is known as Hensen's node, and in amphibians it is known as the Spemann-Mangold organizer. It is induced by the Nieuwkoop center in amphibians, or by the posterior marginal zone in amniotes including birds.
Compartments can be simply defined as separate, different, adjacent cell populations, which upon juxtaposition, create a lineage boundary. This boundary prevents cell movement from cells from different lineages across this barrier, restricting them to their compartment. Subdivisions are established by morphogen gradients and maintained by local cell-cell interactions, providing functional units with domains of different regulatory genes, which give rise to distinct fates. Compartment boundaries are found across species. In the hindbrain of vertebrate embryos, rhombomeres are compartments of common lineage outlined by expression of Hox genes. In invertebrates, the wing imaginal disc of Drosophila provides an excellent model for the study of compartments. Although other tissues, such as the abdomen, and even other imaginal discs are compartmentalized, much of our understanding of key concepts and molecular mechanisms involved in compartment boundaries has been derived from experimentation in the wing disc of the fruit fly.
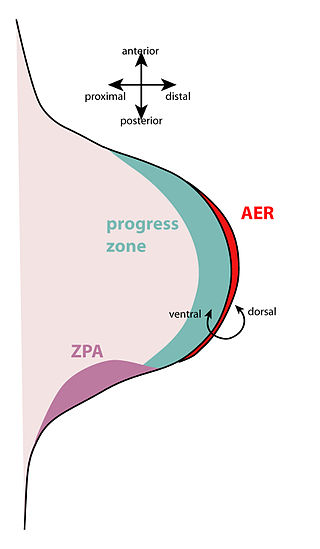
The apical ectodermal ridge (AER) is a structure that forms from the ectodermal cells at the distal end of each limb bud and acts as a major signaling center to ensure proper development of a limb. After the limb bud induces AER formation, the AER and limb mesenchyme—including the zone of polarizing activity (ZPA)—continue to communicate with each other to direct further limb development.
The limb bud is a structure formed early in vertebrate limb development. As a result of interactions between the ectoderm and underlying mesoderm, formation occurs roughly around the fourth week of development. In the development of the human embryo the upper limb bud appears in the third week and the lower limb bud appears four days later.
In the field of developmental biology, regional differentiation is the process by which different areas are identified in the development of the early embryo. The process by which the cells become specified differs between organisms.
The development of the nervous system in humans, or neural development or neurodevelopment involves the studies of embryology, developmental biology, and neuroscience to describe the cellular and molecular mechanisms by which the complex nervous system forms in humans, develops during prenatal development, and continues to develop postnatally.

Homeobox protein OTX2 is a protein that in humans is encoded by the OTX2 gene.
The zone of polarizing activity (ZPA) is an area of mesenchyme that contains signals which instruct the developing limb bud to form along the anterior/posterior axis. Limb bud is undifferentiated mesenchyme enclosed by an ectoderm covering. Eventually, the limb bud develops into bones, tendons, muscles and joints. Limb bud development relies not only on the ZPA, but also many different genes, signals, and a unique region of ectoderm called the apical ectodermal ridge (AER). Research by Saunders and Gasseling in 1948 identified the AER and its subsequent involvement in proximal distal outgrowth. Twenty years later, the same group did transplantation studies in chick limb bud and identified the ZPA. It wasn't until 1993 that Todt and Fallon showed that the AER and ZPA are dependent on each other.

Andrew Gino Lumsden is an English neurobiologist, Emeritus Professor of the University of London and founder in 2000 of the Medical Research Council Centre for Developmental Neurobiology at King's College London.
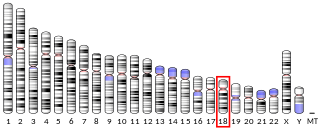
Retinal homeobox protein Rx also known as retina and anterior neural fold homeobox is a protein that in humans is encoded by the RAX gene. The RAX gene is located on chromosome 18 in humans, mice, and rats.
The isthmic organizer, or isthmus organizer, also known as the midbrain−hindbrain boundary (MHB), is a secondary organizer region that develops at the junction of the midbrain and metencephalon. The MHB expresses signaling molecules that regulate the differentiation and patterning of the adjacent neuroepithelium. This allows for the development of the midbrain and hindbrain as well as the specification of neuronal subtypes in these regions. The fact that the MHB is sufficient for the development of the mid and hindbrain was shown in an experiment where quail MHB cells transplanted into the forebrain of a chick were able to induce an ectopic midbrain and cerebellum.


