Long-term memory (LTM) is the stage of the Atkinson–Shiffrin memory model in which informative knowledge is held indefinitely. It is defined in contrast to short-term and working memory, which persist for only about 18 to 30 seconds. Long-term memory is commonly labelled as explicit memory (declarative), as well as episodic memory, semantic memory, autobiographical memory, and implicit memory.

In neuroscience, long-term potentiation (LTP) is a persistent strengthening of synapses based on recent patterns of activity. These are patterns of synaptic activity that produce a long-lasting increase in signal transmission between two neurons. The opposite of LTP is long-term depression, which produces a long-lasting decrease in synaptic strength.

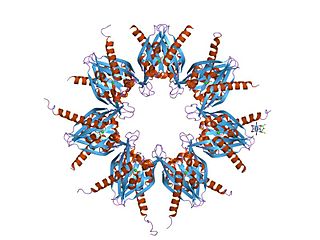
The α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor is an ionotropic transmembrane receptor for glutamate (iGluR) that mediates fast synaptic transmission in the central nervous system (CNS). It has been traditionally classified as a non-NMDA-type receptor, along with the kainate receptor. Its name is derived from its ability to be activated by the artificial glutamate analog AMPA. The receptor was first named the "quisqualate receptor" by Watkins and colleagues after a naturally occurring agonist quisqualate and was only later given the label "AMPA receptor" after the selective agonist developed by Tage Honore and colleagues at the Royal Danish School of Pharmacy in Copenhagen. The GRIA2-encoded AMPA receptor ligand binding core was the first glutamate receptor ion channel domain to be crystallized.
In neuroscience, synaptic plasticity is the ability of synapses to strengthen or weaken over time, in response to increases or decreases in their activity. Since memories are postulated to be represented by vastly interconnected neural circuits in the brain, synaptic plasticity is one of the important neurochemical foundations of learning and memory.
In neurophysiology, long-term depression (LTD) is an activity-dependent reduction in the efficacy of neuronal synapses lasting hours or longer following a long patterned stimulus. LTD occurs in many areas of the CNS with varying mechanisms depending upon brain region and developmental progress.

Brain-derived neurotrophic factor (BDNF), or abrineurin, is a protein that, in humans, is encoded by the BDNF gene. BDNF is a member of the neurotrophin family of growth factors, which are related to the canonical nerve growth factor. Neurotrophic factors are found in the brain and the periphery. BDNF was first isolated from a pig brain in 1982 by Yves-Alain Barde and Hans Thoenen.

Anisomycin, also known as flagecidin, is an antibiotic produced by Streptomyces griseolus which inhibits eukaryotic protein synthesis. Partial inhibition of DNA synthesis occurs at anisomycin concentrations that effect 95% inhibition of protein synthesis. Anisomycin can activate stress-activated protein kinases, MAP kinase and other signal transduction pathways.
Schaffer collaterals are axon collaterals given off by CA3 pyramidal cells in the hippocampus. These collaterals project to area CA1 of the hippocampus and are an integral part of memory formation and the emotional network of the Papez circuit, and of the hippocampal trisynaptic loop. It is one of the most studied synapses in the world and named after the Hungarian anatomist-neurologist Károly Schaffer.
Metaplasticity is a term originally coined by W.C. Abraham and M.F. Bear to refer to the plasticity of synaptic plasticity. Until that time synaptic plasticity had referred to the plastic nature of individual synapses. However this new form referred to the plasticity of the plasticity itself, thus the term meta-plasticity. The idea is that the synapse's previous history of activity determines its current plasticity. This may play a role in some of the underlying mechanisms thought to be important in memory and learning such as long-term potentiation (LTP), long-term depression (LTD) and so forth. These mechanisms depend on current synaptic "state", as set by ongoing extrinsic influences such as the level of synaptic inhibition, the activity of modulatory afferents such as catecholamines, and the pool of hormones affecting the synapses under study. Recently, it has become clear that the prior history of synaptic activity is an additional variable that influences the synaptic state, and thereby the degree, of LTP or LTD produced by a given experimental protocol. In a sense, then, synaptic plasticity is governed by an activity-dependent plasticity of the synaptic state; such plasticity of synaptic plasticity has been termed metaplasticity. There is little known about metaplasticity, and there is much research currently underway on the subject, despite its difficulty of study, because of its theoretical importance in brain and cognitive science. Most research of this type is done via cultured hippocampus cells or hippocampal slices.
Ca2+
/calmodulin-dependent protein kinase II is a serine/threonine-specific protein kinase that is regulated by the Ca2+
/calmodulin complex. CaMKII is involved in many signaling cascades and is thought to be an important mediator of learning and memory. CaMKII is also necessary for Ca2+
homeostasis and reuptake in cardiomyocytes, chloride transport in epithelia, positive T-cell selection, and CD8 T-cell activation.

Protein kinase C, zeta (PKCζ), also known as PRKCZ, is a protein in humans that is encoded by the PRKCZ gene. The PRKCZ gene encodes at least two alternative transcripts, the full-length PKCζ and an N-terminal truncated form PKMζ. PKMζ is thought to be responsible for maintaining long-term memories in the brain. The importance of PKCζ in the creation and maintenance of long-term potentiation was first described by Todd Sacktor and his colleagues at the SUNY Downstate Medical Center in 1993.
Trk receptors are a family of tyrosine kinases that regulates synaptic strength and plasticity in the mammalian nervous system. Trk receptors affect neuronal survival and differentiation through several signaling cascades. However, the activation of these receptors also has significant effects on functional properties of neurons.
Memory consolidation is a category of processes that stabilize a memory trace after its initial acquisition. A memory trace is a change in the nervous system caused by memorizing something. Consolidation is distinguished into two specific processes. The first, synaptic consolidation, which is thought to correspond to late-phase long-term potentiation, occurs on a small scale in the synaptic connections and neural circuits within the first few hours after learning. The second process is systems consolidation, occurring on a much larger scale in the brain, rendering hippocampus-dependent memories independent of the hippocampus over a period of weeks to years. Recently, a third process has become the focus of research, reconsolidation, in which previously consolidated memories can be made labile again through reactivation of the memory trace.

Activity-regulated cytoskeleton-associated protein is a plasticity protein that in humans is encoded by the ARC gene. It was first characterized in 1995. ARC is a member of the immediate-early gene (IEG) family, a rapidly activated class of genes functionally defined by their ability to be transcribed in the presence of protein synthesis inhibitors. ARC mRNA is localized to activated synaptic sites in an NMDA receptor-dependent manner, where the newly translated protein is believed to play a critical role in learning and memory-related molecular processes. Arc protein is widely considered to be important in neurobiology because of its activity regulation, localization, and utility as a marker for plastic changes in the brain. Dysfunction in the production of Arc protein has been implicated as an important factor in understanding various neurological conditions, including amnesia, Alzheimer's disease, Autism spectrum disorders, and Fragile X syndrome. Along with other IEGs such as ZNF268 and HOMER1, ARC is also a significant tool for systems neuroscience as illustrated by the development of the cellular compartment analysis of temporal activity by fluorescence in situ hybridization, or catFISH technique.
Plasticity Product is a term coined by Jerry Rudy to refer to mRNA genetic artifacts and protein products triggered by transcription factors leading to long-lasting long term potentiation.
Synaptic tagging, or the synaptic tagging hypothesis, was first proposed in 1997 by Uwe Frey and Richard G. Morris; it seeks to explain how neural signaling at a particular synapse creates a target for subsequent plasticity-related product (PRP) trafficking essential for sustained LTP and LTD. Although the molecular identity of the tags remains unknown, it has been established that they form as a result of high or low frequency stimulation, interact with incoming PRPs, and have a limited lifespan.
Long-term potentiation (LTP), thought to be the cellular basis for learning and memory, involves a specific signal transmission process that underlies synaptic plasticity. Among the many mechanisms responsible for the maintenance of synaptic plasticity is the cadherin–catenin complex. By forming complexes with intracellular catenin proteins, neural cadherins (N-cadherins) serve as a link between synaptic activity and synaptic plasticity, and play important roles in the processes of learning and memory.

Homosynaptic plasticity is one type of synaptic plasticity. Homosynaptic plasticity is input-specific, meaning changes in synapse strength occur only at post-synaptic targets specifically stimulated by a pre-synaptic target. Therefore, the spread of the signal from the pre-synaptic cell is localized.

Min Zhuo is a pain neuroscientist at the University of Toronto in Canada. He is also the Michael Smith Chair in Neuroscience and Mental Health as well as the Canada Research Chair in Pain and Cognition and a Fellow of the Royal Society of Canada. Supported by the Heidelberg Pain Consortium, Zhou was hosted in 2017-2018 as a Guest Professor at the Pharmacology Institute at Heidelberg University, Heidelberg.
Early long-term potentiation (E-LTP) is the first phase of long-term potentiation (LTP), a well-studied form of synaptic plasticity, and consists of an increase in synaptic strength. LTP could be produced by repetitive stimulation of the presynaptic terminals, and it is believed to play a role in memory function in the hippocampus, amygdala and other cortical brain structures in mammals.