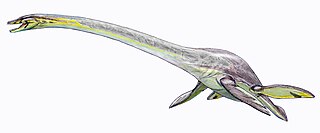
Plesiosauroidea is an extinct clade of carnivorous marine reptiles. They have the snake-like longest neck to body ratio of any reptile. Plesiosauroids are known from the Jurassic and Cretaceous periods. After their discovery, some plesiosauroids were said to have resembled "a snake threaded through the shell of a turtle", although they had no shell.

Elasmosaurus is a genus of plesiosaur that lived in North America during the Campanian stage of the Late Cretaceous period, about 80.5 million years ago. The first specimen was discovered in 1867 near Fort Wallace, Kansas, US, and was sent to the American paleontologist Edward Drinker Cope, who named it E. platyurus in 1868. The generic name means "thin-plate reptile", and the specific name means "flat-tailed". Cope originally reconstructed the skeleton of Elasmosaurus with the skull at the end of the tail, an error which was made light of by the paleontologist Othniel Charles Marsh, and became part of their "Bone Wars" rivalry. Only one incomplete Elasmosaurus skeleton is definitely known, consisting of a fragmentary skull, the spine, and the pectoral and pelvic girdles, and a single species is recognized today; other species are now considered invalid or have been moved to other genera.

The Plesiosauria or plesiosaurs are an order or clade of extinct Mesozoic marine reptiles, belonging to the Sauropterygia.

Umoonasaurus is an extinct genus of plesiosaur belonging to the family Leptocleididae. This genus lived approximately 115 million years ago during the Early Cretaceous period, in shallow seas covering parts of what is now Australia. It was a relatively small animal around 2.5 metres (8.2 ft) long. An identifying trait of Umoonasaurus is three crest-ridges on its skull.

Plesiopleurodon is an extinct genus of Mesozoic marine reptiles, belonging to Sauropterygia, known from the Late Cretaceous of North America. It was named by Kenneth Carpenter based on a complete skull with a mandible, cervical vertebra, and a coracoid. In naming the specimen, Carpenter noted "Of all known pliosaurs, Plesiopleurodon wellesi most closely resembles Liopleurodon ferox from the Oxfordian of Europe, hence the generic reference." It was initially described as a pliosaur due to it short neck, a common trait of the family, although it is in the order Plesiosauria. However, later exploration into the relationships of both orders indicate that not all pliosaurs have short necks and not all plesiosaurs have long necks.

Polycotylidae is a family of plesiosaurs from the Cretaceous, a sister group to Leptocleididae. Polycotylids first appeared during the Albian stage of the Early Cretaceous, before becoming abundant and widespread during the early Late Cretaceous. Several species survived into the final stage of the Cretaceous, the Maastrichtian.

The Bearpaw Formation, also called the Bearpaw Shale, is a geologic formation of Late Cretaceous (Campanian) age. It outcrops in the U.S. state of Montana, as well as the Canadian provinces of Alberta and Saskatchewan, and was named for the Bear Paw Mountains in Montana. It includes a wide range of marine fossils, as well as the remains of a few dinosaurs. It is known for its fossil ammonites, some of which are mined in Alberta to produce the organic gemstone ammolite.
Tuarangisaurus is an extinct genus of elasmosaurid known from New Zealand. The type and only known species is Tuarangisaurus keyesi, named by Wiffen and Moisley in 1986.

Styxosaurus is a genus of plesiosaur of the family Elasmosauridae. Styxosaurus lived during the Campanian age of the Cretaceous period. Three species are known: S. snowii, S. browni, and S. rezaci.

Aristonectes is an extinct genus of large elasmosaurid plesiosaurs that lived during the Maastrichtian stage of the Late Cretaceous. Two species are known, A. parvidens and A. quiriquinensis, whose fossil remains were discovered in what are now Patagonia and Antarctica. Throughout the 20th century, Aristonectes was a difficult animal for scientists to analyze due to poor fossil preparation, its relationships to other genera were uncertain. After subsequent revisions and discoveries carried out from the beginning of the 21st century, Aristonectes is now recognised as the type genus of the subfamily Aristonectinae, a lineage of elasmosaurids characterized by an enlarged skull and a reduced length of the neck.
Kaiwhekea is an extinct genus of plesiosaur from the Late Cretaceous of what is now New Zealand.
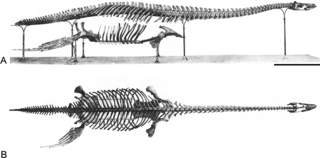
Brancasaurus is a genus of plesiosaur which lived in a freshwater lake in the Early Cretaceous of what is now North Rhine-Westphalia, Germany. With a long neck possessing vertebrae bearing distinctively-shaped "shark fin"-shaped neural spines, and a relatively small and pointed head, Brancasaurus is superficially similar to Elasmosaurus, albeit smaller in size at 3.26 metres (10.7 ft) in length as a subadult.
Eromangasaurus is an extinct genus of elasmosaurid known from northern Queensland of Australia.

Rhomaleosauridae is a family of plesiosaurs from the Earliest Jurassic to the latest Middle Jurassic of Europe, North America, South America and possibly Asia. Most rhomaleosaurids are known from England, many specifically from lower Blue Lias deposits that date back to the earliest Jurassic, just at the boundary with the Triassic. In fact, to date only two undisputed rhomaleosaurids were named from outside Europe - the closely related Borealonectes russelli and Maresaurus coccai from Canada and Argentina, respectively. These two species are also the only Middle Jurassic representatives of the family. Rhomaleosauridae was formally named by Kuhn in 1961, originally proposed to include Rhomaleosaurus cramptoni and its relatives, which have short necks and large heads relatively to plesiosauroids like Elasmosaurus and Plesiosaurus, but longer necks and smaller heads relatively to advanced pliosaurids like Pliosaurus and Kronosaurus.

This timeline of plesiosaur research is a chronologically ordered list of important fossil discoveries, controversies of interpretation, taxonomic revisions, and cultural portrayals of plesiosaurs, an order of marine reptiles that flourished during the Mesozoic Era. The first scientifically documented plesiosaur fossils were discovered during the early 19th century by Mary Anning. Plesiosaurs were actually discovered and described before dinosaurs. They were also among the first animals to be featured in artistic reconstructions of the ancient world, and therefore among the earliest prehistoric creatures to attract the attention of the lay public. Plesiosaurs were originally thought to be a kind of primitive transitional form between marine life and terrestrial reptiles. However, now plesiosaurs are recognized as highly derived marine reptiles descended from terrestrial ancestors.
Vegasaurus is an extinct genus of elasmosaurid plesiosaur known from the Late Cretaceous Snow Hill Island Formation of Vega Island, Antarctic Peninsula. It contains a single species, Vegasaurus molyi.

Kawanectes is a genus of elasmosaurid plesiosaur, a type of long-necked marine reptile, that lived in the marginal marine environment of Late Cretaceous Patagonia. It contains one species, K. lafquenianum, described in 2016 by O'Gorman.

Nakonanectes bradti is an elasmosaurid plesiosaur of the late Cretaceous found in 2010 the state of Montana in the United States. It is one of the most recently known elasmosaurids to have lived in North America. Unlike other elasmosaurids, it has a relatively short neck.








