B cells, also known as B lymphocytes, are a type of white blood cell of the lymphocyte subtype. They function in the humoral immunity component of the adaptive immune system. B cells produce antibody molecules which may be either secreted or inserted into the plasma membrane where they serve as a part of B-cell receptors. When a naïve or memory B cell is activated by an antigen, it proliferates and differentiates into an antibody-secreting effector cell, known as a plasmablast or plasma cell. In addition, B cells present antigens and secrete cytokines. In mammals, B cells mature in the bone marrow, which is at the core of most bones. In birds, B cells mature in the bursa of Fabricius, a lymphoid organ where they were first discovered by Chang and Glick, which is why the B stands for bursa and not bone marrow, as commonly believed.

The T helper cells (Th cells), also known as CD4+ cells or CD4-positive cells, are a type of T cell that play an important role in the adaptive immune system. They aid the activity of other immune cells by releasing cytokines. They are considered essential in B cell antibody class switching, breaking cross-tolerance in dendritic cells, in the activation and growth of cytotoxic T cells, and in maximizing bactericidal activity of phagocytes such as macrophages and neutrophils. CD4+ cells are mature Th cells that express the surface protein CD4. Genetic variation in regulatory elements expressed by CD4+ cells determines susceptibility to a broad class of autoimmune diseases.

In immunology, a memory B cell (MBC) is a type of B lymphocyte that forms part of the adaptive immune system. These cells develop within germinal centers of the secondary lymphoid organs. Memory B cells circulate in the blood stream in a quiescent state, sometimes for decades. Their function is to memorize the characteristics of the antigen that activated their parent B cell during initial infection such that if the memory B cell later encounters the same antigen, it triggers an accelerated and robust secondary immune response. Memory B cells have B cell receptors (BCRs) on their cell membrane, identical to the one on their parent cell, that allow them to recognize antigen and mount a specific antibody response.

In immunology, an Fc receptor is a protein found on the surface of certain cells – including, among others, B lymphocytes, follicular dendritic cells, natural killer cells, macrophages, neutrophils, eosinophils, basophils, human platelets, and mast cells – that contribute to the protective functions of the immune system. Its name is derived from its binding specificity for a part of an antibody known as the Fc region. Fc receptors bind to antibodies that are attached to infected cells or invading pathogens. Their activity stimulates phagocytic or cytotoxic cells to destroy microbes, or infected cells by antibody-mediated phagocytosis or antibody-dependent cell-mediated cytotoxicity. Some viruses such as flaviviruses use Fc receptors to help them infect cells, by a mechanism known as antibody-dependent enhancement of infection.

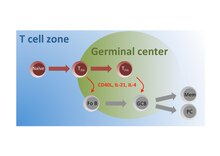
Germinal centers or germinal centres (GCs) are transiently formed structures within B cell zone (follicles) in secondary lymphoid organs – lymph nodes, ileal Peyer's patches, and the spleen – where mature B cells are activated, proliferate, differentiate, and mutate their antibody genes during a normal immune response; most of the germinal center B cells (BGC) are removed by tingible body macrophages. There are several key differences between naive B cells and GC B cells, including level of proliferative activity, size, metabolic activity and energy production. The B cells develop dynamically after the activation of follicular B cells by T-dependent antigen. The initiation of germinal center formation involves the interaction between B and T cells in the interfollicular area of the lymph node, CD40-CD40L ligation, NF-kB signaling and expression of IRF4 and BCL6.

CD154, also called CD40 ligand or CD40L, is a protein that is primarily expressed on activated T cells and is a member of the TNF superfamily of molecules. It binds to CD40 on antigen-presenting cells (APC), which leads to many effects depending on the target cell type. In total CD40L has three binding partners: CD40, α5β1 integrin and integrin αIIbβ3. CD154 acts as a costimulatory molecule and is particularly important on a subset of T cells called T follicular helper cells. On TFH cells, CD154 promotes B cell maturation and function by engaging CD40 on the B cell surface and therefore facilitating cell-cell communication. A defect in this gene results in an inability to undergo immunoglobulin class switching and is associated with hyper IgM syndrome. Absence of CD154 also stops the formation of germinal centers and therefore prohibiting antibody affinity maturation, an important process in the adaptive immune system.

Complement receptor type 2 (CR2), also known as complement C3d receptor, Epstein-Barr virus receptor, and CD21, is a protein that in humans is encoded by the CR2 gene.
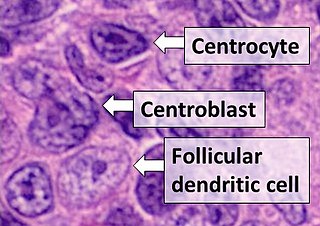
Follicular dendritic cells (FDC) are cells of the immune system found in primary and secondary lymph follicles of the B cell areas of the lymphoid tissue. Unlike dendritic cells (DC), FDCs are not derived from the bone-marrow hematopoietic stem cell, but are of mesenchymal origin. Possible functions of FDC include: organizing lymphoid tissue's cells and microarchitecture, capturing antigen to support B cell, promoting debris removal from germinal centers, and protecting against autoimmunity. Disease processes that FDC may contribute include primary FDC-tumor, chronic inflammatory conditions, HIV-1 infection development, and neuroinvasive scrapie.

Interleukin 21 (IL-21) is a protein that in humans is encoded by the IL21 gene.

Chemokine ligand 18 (CCL18) is a small cytokine belonging to the CC chemokine family. The functions of CCL18 have been well studied in laboratory settings, however the physiological effects of the molecule in living organisms have been difficult to characterize because there is no similar protein in rodents that can be studied. The receptor for CCL18 has been identified in humans only recently, which will help scientists understand the molecule's role in the body.
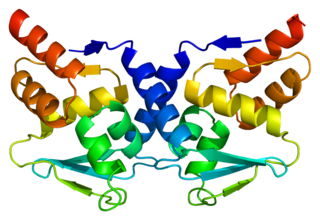
Bcl-6 is a protein that in humans is encoded by the BCL6 gene. BCL6 is a master transcription factor for regulation of T follicular helper cells proliferation. BCL6 has three evolutionary conserved structural domains. The interaction of these domains with corepressors allows for germinal center development and leads to B cell proliferation.

ICOS ligand is a protein that in humans is encoded by the ICOSLG gene located at chromosome 21. ICOSLG has also been designated as CD275.

CD160 antigen is a protein that in humans is encoded by the CD160 gene.

HLA class II histocompatibility antigen, DX beta chain is a protein that in humans is encoded by the HLA-DQB2 gene.

Marginal-zone B cells are noncirculating mature B cells that in humans segregate anatomically into the marginal zone (MZ) of the spleen and certain other types of lymphoid tissue. The MZ B cells within this region typically express low-affinity polyreactive B-cell receptors (BCR), high levels of IgM, Toll-like receptors (TLRs), CD21, CD1, CD9, CD27 with low to negligible levels of secreted-IgD, CD23, CD5, and CD11b that help to distinguish them phenotypically from follicular (FO) B cells and B1 B cells.
Within the immune system, Follicular B cells are a type of B cell that reside in primary and secondary lymphoid follicles of secondary and tertiary lymphoid organs, including spleen and lymph nodes. Antibody responses against proteins are believed to involve follicular B cell pathways in secondary lymphoid organs.

A centroblast generally refers to an activated B cell that is enlarged and is rapidly proliferating in the germinal center of a lymphoid follicle. They are specifically located in the dark zone of the germinal center. Centroblasts form from naive B cells being exposed to follicular dendritic cell cytokines, such as IL-6, IL-15, 8D6, and BAFF. Stimulation from helper T cells is also required for centroblast development. Interaction between CD40 ligand on an activated T helper cell and the B cell CD40 receptor induces centroblasts to express activation-induced cytidine deaminase, leading to somatic hypermutation, allowing the B cell receptor to potentially gain stronger affinity for an antigen. In the absence of FDC and helper T cell stimulation, centroblasts are unable to differentiate and will undergo CD95-mediated apoptosis.
Regulatory B cells (Bregs or Breg cells) represent a small population of B cells that participates in immunomodulation and in the suppression of immune responses. The population of Bregs can be further separated into different human or murine subsets such as B10 cells, marginal zone B cells, Br1 cells, GrB+B cells, CD9+ B cells, and even some plasmablasts or plasma cells. Bregs regulate the immune system by different mechanisms. One of the main mechanisms is the production of anti-inflammatory cytokines such as interleukin 10 (IL-10), IL-35, or transforming growth factor beta (TGF-β). Another known mechanism is the production of cytotoxic Granzyme B. Bregs also express various inhibitory surface markers such as programmed death-ligand 1 (PD-L1), CD39, CD73, and aryl hydrocarbon receptor. The regulatory effects of Bregs were described in various models of inflammation, autoimmune diseases, transplantation reactions, and in anti-tumor immunity.
T-cell receptor revision is a process in the peripheral immune system which is used by mature T cells to alter their original antigenic specificity based on rearranged T cell receptors (TCR). This process can lead either to continuous appearance of potentially self-reactive T cells in the body, not controlled by the central tolerance mechanism in the thymus or better eliminate such self-reactive T cells on the other hand and thus contributing to peripheral tolerance - the extent of each has not been completely understood yet. This process occurs during follicular helper T cell formation in lymph node germinal centers.

Elina Zúñiga is an Argentinian Immunologist and Professor of Molecular Biology in the Division of Biological Sciences at the University of California, San Diego. Zúñiga has made critical discoveries regarding host-virus interactions in both acute and chronic infections. Using lymphocytic choriomeningitis models (LCMV) and murine cytomegalovirus models, her laboratory at UCSD studies host immune adaptations in chronic viral disease and methods of viral suppression of the immune system in order to develop novel methods and identify novel targets of anti-viral defence. In 2018, Zúñiga was chosen by the American Association of Immunologists to give the international Vanguard Lecture. Zúñiga is also co-founder of the Global Immunotalks series which she and Carla Rothlin started in 2020 as a means to make cutting-edge immunology research freely available and easily accessible to a global audience.