Types of heteroplasmy
In order for heteroplasmy to occur, organelles must contain a genome and, in turn, a genotype. In animals, mitochondria are the only organelles that contain their own genomes, so these organisms will only have mitochondrial heteroplasmy. In contrast, photosynthetic plants contain mitochondria and chloroplasts, each of which contains plastid genomes. Therefore, plant heteroplasmy occurs in two dimensions. [4]
Microheteroplasmy
Microheteroplasmy is the presence of mutations levels of up to about 2−5% of mitochondrial genomes. In human mitochondrial DNA, microheteroplasmy constitutes hundreds of independent mutations in one organism, with each mutation usually found in 1–2% of all mitochondrial genomes. [5]
The distinction of microheteroplasmy and more gross heteroplasmy is dictated by technical considerations - classical DNA sequencing of mitochondrial DNA by the use of PCR is capable only of detecting mutations at levels of 10% or more, as a result of which mutations at lower levels were never systematically observed until the work of Lin et al. [6]
As it became apparent after the use of Lin's cloning and sequencing strategy, capable of detecting mutations at levels of 1% or less, such low-level heteroplasmy, or microheteroplasmy, is exceedingly common, and is in fact the most common form of mutational damage to human DNA found to date. In aged adults, each copy of mitochondrial DNA has on average 3.3 mutations changing protein structure. This exceeds previous estimates by more than three orders of magnitude.
The discovery of microheteroplasmy lends support to the mitochondrial theory of aging, and has already been linked to the causation of Parkinson's disease. [7]

A mitochondrion is an organelle found in the cells of most eukaryotes, such as animals, plants and fungi. Mitochondria have a double membrane structure and use aerobic respiration to generate adenosine triphosphate (ATP), which is used throughout the cell as a source of chemical energy. They were discovered by Albert von Kölliker in 1857 in the voluntary muscles of insects. The term mitochondrion was coined by Carl Benda in 1898. The mitochondrion is popularly nicknamed the "powerhouse of the cell", a phrase coined by Philip Siekevitz in a 1957 article of the same name.
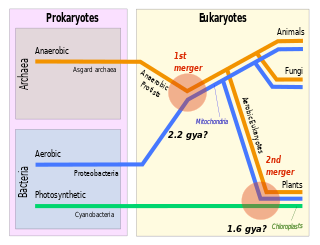
Symbiogenesis is the leading evolutionary theory of the origin of eukaryotic cells from prokaryotic organisms. The theory holds that mitochondria, plastids such as chloroplasts, and possibly other organelles of eukaryotic cells are descended from formerly free-living prokaryotes taken one inside the other in endosymbiosis. Mitochondria appear to be phylogenetically related to Rickettsiales bacteria, while chloroplasts are thought to be related to cyanobacteria.

In genetics, dominance is the phenomenon of one variant (allele) of a gene on a chromosome masking or overriding the effect of a different variant of the same gene on the other copy of the chromosome. The first variant is termed dominant and the second is called recessive. This state of having two different variants of the same gene on each chromosome is originally caused by a mutation in one of the genes, either new or inherited. The terms autosomal dominant or autosomal recessive are used to describe gene variants on non-sex chromosomes (autosomes) and their associated traits, while those on sex chromosomes (allosomes) are termed X-linked dominant, X-linked recessive or Y-linked; these have an inheritance and presentation pattern that depends on the sex of both the parent and the child. Since there is only one copy of the Y chromosome, Y-linked traits cannot be dominant or recessive. Additionally, there are other forms of dominance, such as incomplete dominance, in which a gene variant has a partial effect compared to when it is present on both chromosomes, and co-dominance, in which different variants on each chromosome both show their associated traits.

Mitochondrial DNA is the DNA located in mitochondria, cellular organelles within eukaryotic cells that convert chemical energy from food into a form that cells can use, such as adenosine triphosphate (ATP). Mitochondrial DNA is only a small portion of the DNA in a eukaryotic cell; most of the DNA can be found in the cell nucleus and, in plants and algae, also in plastids such as chloroplasts.

Leigh syndrome is an inherited neurometabolic disorder that affects the central nervous system. It is named after Archibald Denis Leigh, a British neuropsychiatrist who first described the condition in 1951. Normal levels of thiamine, thiamine monophosphate, and thiamine diphosphate are commonly found, but there is a reduced or absent level of thiamine triphosphate. This is thought to be caused by a blockage in the enzyme thiamine-diphosphate kinase, and therefore treatment in some patients would be to take thiamine triphosphate daily.

Homoplasmy is a term used in genetics to describe a eukaryotic cell whose copies of mitochondrial DNA are all identical. In normal and healthy tissues, all cells are homoplasmic. Homoplasmic mitochondrial DNA copies may be normal or mutated; however, most mutations are heteroplasmic. It has been discovered, though, that homoplasmic mitochondrial DNA mutations may be found in human tumors.

Human mitochondrial genetics is the study of the genetics of human mitochondrial DNA. The human mitochondrial genome is the entirety of hereditary information contained in human mitochondria. Mitochondria are small structures in cells that generate energy for the cell to use, and are hence referred to as the "powerhouses" of the cell.

Non-Mendelian inheritance is any pattern in which traits do not segregate in accordance with Mendel's laws. These laws describe the inheritance of traits linked to single genes on chromosomes in the nucleus. In Mendelian inheritance, each parent contributes one of two possible alleles for a trait. If the genotypes of both parents in a genetic cross are known, Mendel's laws can be used to determine the distribution of phenotypes expected for the population of offspring. There are several situations in which the proportions of phenotypes observed in the progeny do not match the predicted values.
Genetics, a discipline of biology, is the science of heredity and variation in living organisms.
petite (ρ–) is a mutant first discovered in the yeast Saccharomyces cerevisiae. Due to the defect in the respiratory chain, 'petite' yeast are unable to grow on media containing only non-fermentable carbon sources and form small colonies when grown in the presence of fermentable carbon sources. The petite phenotype can be caused by the absence of, or mutations in, mitochondrial DNA, or by mutations in nuclear-encoded genes involved in oxidative phosphorylation. A neutral petite produces all wild type progeny when crossed with wild type.

A nuclear gene is a gene whose physical DNA nucleotide sequence is located in the cell nucleus of a eukaryote. The term is used to distinguish nuclear genes from genes found in mitochondria or chloroplasts. The vast majority of genes in eukaryotes are nuclear.
Extrachromosomal DNA is any DNA that is found off the chromosomes, either inside or outside the nucleus of a cell. Most DNA in an individual genome is found in chromosomes contained in the nucleus. Multiple forms of extrachromosomal DNA exist, and, while some of these serve important biological functions, they can also play a role in diseases such as cancer.
In genetics, paternal mtDNA transmission and paternal mtDNA inheritance refer to the incidence of mitochondrial DNA (mtDNA) being passed from a father to his offspring. Paternal mtDNA inheritance is observed in a small proportion of species; in general, mtDNA is passed unchanged from a mother to her offspring, making it an example of non-Mendelian inheritance. In contrast, mtDNA transmission from both parents occurs regularly in certain bivalves.
Extranuclear inheritance or cytoplasmic inheritance is the transmission of genes that occur outside the nucleus. It is found in most eukaryotes and is commonly known to occur in cytoplasmic organelles such as mitochondria and chloroplasts or from cellular parasites like viruses or bacteria.
NUMT, pronounced "new might", is an acronym for "nuclear mitochondrial DNA" segment or genetic locus coined by evolutionary geneticist, Jose V. Lopez, which describes a transposition of any type of cytoplasmic mitochondrial DNA into the nuclear genome of eukaryotic organisms.
The CoRR hypothesis states that the location of genetic information in cytoplasmic organelles permits regulation of its expression by the reduction-oxidation ("redox") state of its gene products.
Uniparental inheritance is a non-Mendelian form of inheritance that consists of the transmission of genotypes from one parental type to all progeny. That is, all the genes in offspring will originate from only the mother or only the father. This phenomenon is most commonly observed in eukaryotic organelles such as mitochondria and chloroplasts. This is because such organelles contain their own DNA and are capable of independent mitotic replication that does not endure crossing over with the DNA from another parental type. Although uniparental inheritance is the most common form of inheritance in organelles, there is increased evidence of diversity. Some studies found doubly uniparental inheritance (DUI) and biparental transmission to exist in cells. Evidence suggests that even when there is biparental inheritance, crossing-over doesn't always occur. Furthermore, there is evidence that the form of organelle inheritance varied frequently over time. Uniparental inheritance can be divided into multiple subtypes based on the pathway of inheritance.
This glossary of genetics and evolutionary biology is a list of definitions of terms and concepts used in the study of genetics and evolutionary biology, as well as sub-disciplines and related fields, with an emphasis on classical genetics, quantitative genetics, population biology, phylogenetics, speciation, and systematics. Overlapping and related terms can be found in Glossary of cellular and molecular biology, Glossary of ecology, and Glossary of biology.
Biparental inheritance is a type of biological inheritance where the progeny inherits a maternal and a paternal allele for one gene. It is one of the criteria for Mendelian inheritance. Sexual reproduction, where offspring result from the fusion of gametes from two parents, is the most common form of biparental inheritance. While less common, cases of biparental inheritance in extranuclear genes have been documented, such as biparental inheritance of mitochondrial DNA, or chloroplast DNA in plants. Biparental inheritance of nuclear DNA by way of sexual reproduction can allow for new combinations of alleles from each contributing parent. The production of gametes through meiosis can sometimes include recombination, or crossing-over, which is a possibility for novel combinations of alleles.
Professor Patrick Francis Chinnery, FRCP, FRCPath, FMedSci, is a neurologist, clinician scientist, and Wellcome Trust Principal Research Fellow based in the Medical Research Council Mitochondrial Biology Unit and the University of Cambridge, where he is also Professor of Neurology and Head of the Department of Clinical Neurosciences.

