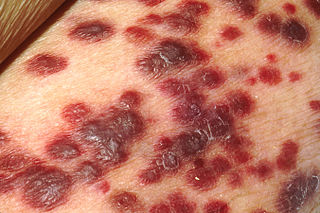
HHV-6A has been described as more neurovirulent,[3] and as such is more frequently found in patients with neuroinflammatory diseases such as multiple sclerosis.[4] HHV-6 (and HHV-7) levels in the brain are also elevated in people with Alzheimer's disease.[5]
HHV-6B primary infection is the cause of the common childhood illness exanthema subitum (also known as roseola infantum or sixth disease). It is passed on from child to child. It is uncommon for adults to contract this disease as most people have had it by kindergarten, and once contracted, immunity arises and prevents future reinfection. Additionally, HHV-6B reactivation is common in transplant recipients, which can cause several clinical manifestations such as encephalitis, bone marrow suppression, and pneumonitis.[6]
A variety of tests are used in the detection of HHV-6, some of which do not differentiate the two species.[7]
Both viruses can cause transplacental infection and be passed on to a newborn.[8]
HHV-6A and Infertility
A 2016 study showed that 43% of women with unexplained infertility tested positive for HHV-6A compared to 0% in the fertile control group. HHV-6A was found present in endometrial epithelial cells from women with unexplained infertility but not in their blood. In the context of infertility, this discovery underscores the importance of targeted testing for HHV-6A within the uterine environment, as the virus was not detected in the bloodstream of the affected individuals. Effective diagnosis, therefore, requires tests that are capable of distinguishing between active and latent HHV-6A infections specifically in endometrial tissue, highlighting the need for tissue-specific viral detection methods in assessing and managing infertility associated with HHV-6A.[9]
A 2018 study found 37% of women experiencing recurrent implantation failure after IVF/ET had HHV-6A in their endometrial biopsies, compared to 0% in control groups.[10]
A 2019 study confirmed the presence of HHV-6A infection in 40% of idiopathic infertile women. Identifying the effect of HHV-6A infection on endometrial immune status opens up a new perspectives on fertility care. It is possible to choose antiviral therapies and non-hormonal approaches for women with unexplained infertility characterized by HHV-6A to increase their pregnancy rate.[11]
Testing for HHV-6
The table below presents a comprehensive overview of various diagnostic tests used to detect human herpesvirus 6 (HHV-6), detailing their ability to distinguish between active and latent infections.[12] It also includes insights on the interpretation of test results, identifies providers that offer these tests, and indicates which methods are suitable for detecting HHV-6A in the endometrial lining—an important consideration for evaluating potential causes of infertility in women. The table serves as a guide for healthcare professionals to select appropriate diagnostic tests for HHV-6.
Diagnostic Tests for HHV-6 Infection and Their Utility in Uterine Environment Analysis
Yes, present only during or shortly after active infection.
Lack of IgM doesn't exclude the possibility of active infection, as chronic infections may not show IgM presence.
IgM tests are widely available.
No
PCR for HHV-6 in Fluids
Yes, presence in fluids suggests an acute infection.
Negative results for HHV-6 DNA don't exclude tissue-based persistent infections. Repeat testing and whole blood analysis are recommended for thorough evaluation.
PCR testing is a common service offered by many laboratories.
No
Whole Blood PCR (Viral Load Count)
Yes, viral load above a certain threshold indicates active infection.
In healthy individuals, viral loads are usually below the detection limit of standard labs.
PCR viral load testing is standard in many diagnostic labs.
No
Whole Blood PCR (Presence/Absence)
No, cannot distinguish between active and latent states.
The majority of healthy individuals will have latent HHV-6B. This test helps identify the type of HHV-6 but not its activity.
Not specified in the original table.
No
Immunohistochemical Testing
Yes, can detect virus replication through protein expression.
Can discern between HHV-6A and HHV-6B activity in fixed tissue.
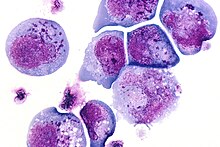
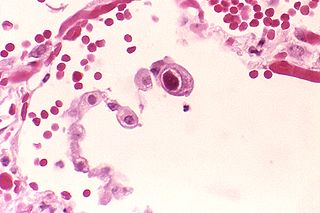
Histological slide of the human herpes virus-6 showing infected cells, with inclusion bodies in both the nucleus and the cytoplasm.
During 1986, Syed Zaki Salahuddin, Dharam Ablashi, and Robert Gallo cultivated peripheral blood mononuclear cells from patients with AIDS and lymphoproliferative illnesses. Short-lived, large, refractile cells that frequently contained intranuclear and/or intracytoplasmic inclusion bodies were documented. Electron microscopy revealed a novel virus that they named human B-lymphotropic virus (HBLV).[13][14]
Shortly after its discovery, Ablashi et al. described five cell lines that can be infected by the newly discovered HBLV. They published that HSB-2, a particular T-cell line, is highly susceptible to infection. Ablashi's pioneering research concluded by suggesting that the virus name be changed from HBLV to HHV-6, in accord with the published provisional classification of herpes viruses.[15][16]
Years later, HHV-6 was divided into subtypes. Early research (1992) described two very similar, yet unique variants: HHV-6A and HHV-6B. The distinction was warranted due to unique restriction endonuclease cleavages, monoclonal antibody reactions,[17] and growth patterns.[18]
HHV-6A includes several adult-derived strains and its disease spectrum is not well defined, although it is thought by some to be more neurovirulent.[19][20] HHV-6B is commonly detected in children with roseola infantum, as it is the etiologic agent for this condition. Within these two viruses is a sequence homology of 95%.[21]
In 2012, HHV-6A and HHV-6B were officially recognized as distinct species.[1]
Herpesvirus was established as a genus in 1971 in the first report of the ICTV. This genus consisted of 23 viruses among 4 groups.[22] In 1976, a second ICTV report was released in which this genus was elevated to the family level— the herpetoviridae. Because of possible confusion with viruses derived from reptiles, the family name was changed in the third report (1979) to herpesviridae. In this report, the family Herpesviridae was divided into 3 subfamilies (alphaherpesvirinae, betaherpesvirinae and gammaherpesvirinae) and 5 unnamed genera; 21 viruses were recognized as members of the family.[23]
In 2009, the order Herpesvirales was created. This was necessitated by the discovery that the herpes viruses of fish and molluscs are only distantly related to those of birds and mammals. Order Herpesvirales contains three families, the Herpesviridae, which contains the long-recognized herpesviruses of mammals, birds, and reptiles, plus two new families— the family Alloherpesviridae which incorporates herpes viruses of bony fish and frogs, and the family Malacoherpesviridae which contains viruses of molluscs.[24]
As of 2012, this order currently has 3 families, 4 subfamilies (1 unassigned), 18 genera (4 unassigned) and 97 species.[1]
Structure
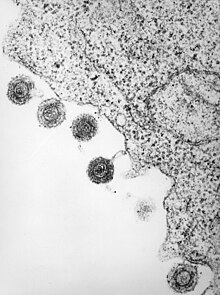
The diameter of an HHV-6 virion is about 2000 angstroms.[14] The virion's outer portion consists of a lipid bilayer membrane that contains viral glycoproteins and is derived from that of the host. Below this membrane envelope is a tegument which surrounds an icosahedralcapsid, composed of 162 capsomeres. The protective capsid of HHV-6 contains double stranded linear DNA.
During maturation of HHV-6 virions, human cell membranes are used to form viral lipid envelopes (as is characteristic of all enveloped viruses). During this process HHV-6 utilizes lipid rafts, which are membranous microdomains enriched by cholesterol, sphingolipids, and glycosylphosphatidylinositol-anchored proteins.[25] Early researchers suspected that HHV-6 virions mature in the nucleus; some even incorrectly published this, as they generalized and applied to HHV-6 what was known about other viruses. However, researched published in 2009 suggests that the HHV-6 virus utilizes trans-Golgi-network-derived vesicles for assembly.[25]
Genome
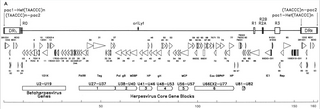
HHV-6B genome from Dominguez et al. 1999
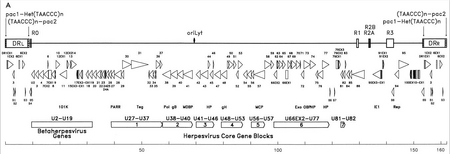
The genetic material of HHV-6 is composed of linear (circular during an active infection), double stranded DNA which contains an origin of replication, two 8–10 kb left and right direct repeat termini, and a unique segment that is 143–145kb.[27]
The origin of replication (often labeled as "oriLyt" in the literature) is where DNA replication begins.[26] The direct repeat termini (DRL and DRR) possess a repeated TTAGGG sequence, identical to that of human telomeres. Variability in the number of telomeric repeats is observed in the range of 15–180.[28][29] These termini also contain pac-1 and pac-2 cleavage and packing signals that are conserved among herpesviruses.
The unique segment contains seven major core gene blocks (U27–U37, U38–U40, U41–U46, U48–U53, U56–U57, U66EX2–U77, and U81–U82),[26] which is also characteristic of herpesviruses. These conserved genes code for proteins that are involved in replication, cleavage, and packing of the viral genome into a mature virion.[28] Additionally, they code for a number of immunomodulatory proteins. The unique segment also possesses a block of genes (U2–U19) that are conserved among HHV-6, HHV-7, and cytomegaloviruses (the betaherpesviruses). A number of the unique segment genes are associated with, for instance, the HCMV US22 family (InterPro:IPR003360). The table below outlines some of their known properties.[26]
Genes
Gene
Stage
Properties
IE-A (IE1? U89?)
Immediate early
Part of IE locus [30] — impairs interferon gene expression to restrict the development of cellular anti-viral measures, favoring a successful infection— not in membrane— activates viral DNA polymerases, involved in rolling circle replication— expression of this gene may be modulated by micro RNAs [31]
IE-B
Immediate early
Part of IE locus [30] Activates viral DNA polymerases, involved in rolling circle replication
DR1
HCMV US22 gene family
DR6
HCMV US22 gene family, transactivator, oncogene
DR7/U1
SR domain, malignant transforming activity, binds to p53
Strongly immunoreactive virion protein [26] — antigenic tegument protein
U12
Chemokine G protein-coupled receptor
U13
CMV: Represses US3 transcription
U14
Binds and incorporates p53 into viral particles— HCMV UL25 gene family— antigenic tegument protein
U15
HCMV UL25 gene family
U17
HCMV UL25 gene family— tegument protein
U18
IE-B
Membrane glycoprotein
U19
IE-B protein
Glycoprotein
U20
Glycoprotein (specific to Roseolovirus) predicted immunoglobulin structure
U21
Binds to MHC-1 molecules and prevents antigen presenting cells from presenting HHV-6 peptides— glycoprotein, downregulates HLA I (specific to Roseolovirus)
U22
Late gene
Glycoprotein (absent from HHV-7, specific to Roseolovirus)
Inhibits proper T cell activation, reducing secretion of cytokines at infection site— phosphorylation target for kinases— glycoprotein M (gM) (specific to Roseolovirus)
U25
HCMV UL22 gene family, tegument protein
U26
Putative multiple transmembrane protein
U27
DNA polymerase processivity factory
U28
Ribonucleotide reductase large subunit, tegument protein
Glycoprotein O, associates with lipid rafts, exists in two forms, gO-120K and gO-80K, and gO-80K contains complex type N-linked oligosaccharides which are incorporated into viral particles
U48 (gH, gp100)
Glycoprotein gH, virion constituent, part of CD46 gQ1/gQ2/gL/gH ligand complex, associates with lipid rafts
U49
Virion-associated regulatory protein, fusion protein
U50
DNA packaging
U51
Early gene
G protein-coupled chemokine receptor, preventing expression greatly reduces replication— increases intracellular levels of second messenger inositol phosphate, promotes chemotaxis– early gene, along with U41 and U69 [7]
Tegument protein kinase (Ganciclovir kinase) involved in replication [30]
U70
Alkaline exonuclease
U71
Myristylated virion protein
U72 (gM)
Glycoprotein M
U73
Origin-binding protein
U74
DNa helicase-primase complex
U75
Tegument protein
U76
DNA packaging, virion protein
U77
Helicase-primase complex
U79
Transcriptional activation
U80
Predicted immunoglobulin structure
U81
Uracil-DNA glycosylase
U82 (gL, gp80)
Glycoprotein L, virion constituent, part of CD46 gQ1/gQ2/gL/gH ligand complex, associates with lipid rafts
U83
Secreted chemotactic (chemoattractant) glycoprotein, binds to chemokine receptors, recruits host cells that secrete chemokines specific to U51
U85
Glycoprotein (specific to Roseolovirus)
U86
IE-2
IE-2 transactivator
U88
IE-A
U90
IE-A (IE 1)
Transactivator
U91
IE-A, Glycoprotein
U94
Latency (immediate early or early gene)
Involved in transcriptional repression of lytic genes– aids in the specific integration of HHV-6A/HHV-6B into the telomeres— highly expressed during latency— parvovirus rep homolog (absent in HHV-7)
U95
CMV US22 gene family– colocalizes and interacts with the mitochondrial GRIM-19 protein, an essential component of the oxidative phosphorylation system [7] — binds to nuclear factor-kappa B (NF-κB), deregulation of which has been postulated to contribute to cancer [19]
U100 (Gp82-105)
Late gene
Glycoprotein Q, virion constituent, associates with lipid rafts
gQ1
Glycoprotein, complexes with gH and gL to form viral ligand to CD46 receptor– modified by N-glycosylation— expressed in two different forms: an 80-kDa form (gQ1-80K) and a 74-kDa form (gQ1-74K) – only gQ1-80K, but not gQ1-74K, forms the CD46 ligand complex with gQ2, gH, and gL [32] Associates with lipid rafts.
gM1
Lipid-raft-specific ganglioside, incorporated into virion
gQ2
Glycoprotein, forms gH/gL/gQ1/gQ2 complex, part of receptor ligand– essential for viral growth, associates with lipid rafts— exists in two forms: gQ2-34K and gQ2-37K
Micro RNAs
hhv6b-miR-Ro6-1, -Ro6-2, -Ro6-3, and -Ro6-4. May regulate early transcription
P100 aka p101
Immunogenic, constituent of tegument
ORF-1 (DR7)
Binds and inhibits transcriptional activity of p53 – can transform human epidermal keratinocytes and NIH 3T3 cells in vitro– cells expressing ORF-1 protein produce fibrosarcomas when injected into nude mice
Viral entry
HHV-6 receptor
When an extracellular HHV-6 virion comes across human cells, it encounters the human receptor protein cluster of differentiation 46 (CD46), which plays a role in regulating the complement system. The CD46 protein possesses a single variable region, as a result of alternative splicing. As such, at least fourteen isoforms of CD46 exist, all of which bind HHV-6a.[33]
The extracellular region of CD46 contains four short consensus repeats of about 60 amino acids that fold into a compact beta-barrel domain surrounded by flexible loops.[28] As has been demonstrated for CD46 with other ligands, the CD46 protein structure linearizes upon binding HHV-6. While their precise interaction has not yet been determined, the second and third SCR domains have been demonstrated as required for HHV-6 receptor binding and cellular entry.
HHV-6 receptor ligand
Mori et al. first identified the gene product gQ1, a glycoprotein unique to HHV-6, and found that it forms a complex with gH and gL glycoproteins.[17][34] They believed that this heterotrimer complex served as the viral ligand for CD46.[27] Soon thereafter, another glycoprotein named gQ2 was identified and found to be part of the gH/gL/gQ1 ligand complex, forming a heterotetramer that was positively identified as the viral CD46 ligand.[34] The exact process of entry is not yet well understood.
Salivary glands
The salivary glands have been described as an in vivo reservoir for HHV-6 infection.[28]
Leukocytes
Researchers[35] conducted a study to show that T cells are highly infectable by HHV-6.
Nervous system
During the year 2011, researchers at the National Institutes of Health attempted to elucidate the then unknown method whereby HHV-6a gains entry into the nervous system. As such, they autopsied the brains of around 150 subjects. When various anatomical regions were assayed for their viral load, olfactory tissues were found to have the highest HHV-6 content. They concluded that these tissues are the entry point for HHV-6a.[21]
The results above are consistent with those of previous studies that involved HSV-1 (and a number of other viruses), which also disseminates into the CNS through olfactory tissue.[36]
Researchers also hypothesized that olfactory ensheathing cells (OECs), a group of specialized glial cells found in the nasal cavity, may have a role in HHV-6 infectivity.[21] They suspected this association as a result of OECs having properties similar to those of astrocytes, another type of glial cell that was previously identified as being susceptible to HHV-6 infection.[37] Research continued by infecting OECs in vitro with both types of HHV-6. Ultimately, only OECs in which HHV-6a was used tested positive for signs of de novo viral synthesis, as is also characteristic of astrocytes.[37]
Cellular activity
Once inside, two outcomes have been described: active and inactive infections.
Active infection
Active infections involve the linear dsDNA genome circularizing by end to end covalent linkages. This process was first reported for the herpes simplex virus.[29] Once circularized, HHV-6 begins to express what are known as "immediate early" genes. These gene products are believed to be transcription activators[7] and may be regulated by the expression of viral micro RNAs.[31] Subsequent expression of "early genes" then occurs and activates, for instance, viral DNA polymerases. Early genes are also involved in the rolling circle replication that follows.[28]
HHV-6's replication results in the formation of concatemers, which are long molecules that contain several repeats of a DNA sequence.[38] These long concatemers are then cleaved between the pac-1 and pac-2 regions for packaging of the genome into individual virions.[29]
Inactive infection
Not all newly infected cells begin rolling circle replication. Herpesviruses may enter a latent stage, inactively infecting their human host. Since its discovery in 1993, this phenomenon has been found among all of the betaherpesviruses.[39]
Other betaherpesviruses establish latency as a nuclear episome, which is a circular DNA molecule (analogous to plasmids). For HHV-6, latency is believed to occur exclusively through the integration of viral telomeric repeats into human subtelomeric regions.[20] Only one other virus, Marek's disease virus, is known to achieve latency in this fashion.[7] This phenomenon is possible as a result of the telomeric repeats found within the direct repeat termini of HHV-6's genome.
The right direct repeat terminus integrates within 5 to 41 human telomere repeats, and preferentially does so into the proximal end[40] of chromosomes 9, 17, 18, 19, and 22, but has also occasionally been found in chromosomes 10 and 11.[38] Nearly 70 million individuals are suspected to carry chromosomally integrated HHV-6.[20][38]
A number of genes expressed by HHV-6 are unique to its inactive latency stage. These genes involve maintaining the genome and avoiding destruction of the host cell.[40] For instance, the U94 protein is believed to repress genes that are involved in cellular lysis (apoptosis) and also may aid in telomeric integration.[28] Once stored in human telomeres, the virus is reactivated intermittently.[40]
Reactivation and transplantation
The specific triggers for reactivation are not well understood. Some researchers have suggested that injury, physical or emotional stress, and hormonal imbalances could be involved.[41]
Researchers during 2011 discovered that reactivation can positively be triggered in vitro by histone deacetylase inhibitors. Once reactivation begins, the rolling circle process is initiated and concatemers are formed as described above.[28]
A study published in The Journal of Infectious Diseases in 2024 investigated the reactivation of inherited chromosomally integrated human herpesvirus 6 (iciHHV-6B) in a liver transplant recipient and its impact on the graft. The research, conducted by Hannolainen et al., used hybrid capture sequencing and various molecular techniques to analyze the viral sequences and host immune response. The findings demonstrated active replication of iciHHV-6B and significant immune activation, suggesting the pathological impact of viral reactivation on transplant outcomes. The study emphasizes the importance of monitoring iciHHV-6 reactivation in transplant patients.[42]
Interactions
Human herpesvirus 6 lives primarily on humans and, while variants of the virus can cause mild to fatal illnesses, can live commensally on its host.[18] It has been demonstrated that HHV-6 fosters the progression of HIV-1 upon coinfection in T cells.[43] HHV-6 upregulates the expression of the primary HIV receptor CD4, thus expanding the range of HIV susceptible cells. Several studies also have shown that HHV-6 infection increases production of inflammatory cytokines that enhance in vitro expression of HIV-1, such as TNF-alpha,[44] IL-1 beta, and IL-8.[45] A more recent in vivo study shows HHV-6A coinfection to dramatically accelerate the progression from HIV to AIDS in pigtailed macaques.[46]
HHV-6 has also been demonstrated to transactivate Epstein–Barr virus.[36]
Epidemiology
Age
Humans acquire the virus at an early age, some as early as less than one month of age. HHV-6 primary infections account for up to 20% of infant emergency room visits for fever in the United States[47][48] and are associated with several more severe complications, such as encephalitis, lymphadenopathy, myocarditis and myelosuppression. The prevalence of the virus in the body increases with age (rates of infection are highest among infant between 6 and 12 months old) and it is hypothesized that this is due to the loss of maternal antibodies in a child that protect him or her from infections.[18]
There are inconsistencies with the correlations between age and seropositivity: According to some reports there is a decrease of seropositivity with the increase of age, while some indicate no significant decline, and others report an increased rate of seropositivity for individuals age 62 and older. After primary infection, latency is established in salivary glands, hematopoietic stem cells, and other cells, and exists for the lifetime of the host.
Geographical distribution
The virus is known to be widespread around the world. An HHV-6 infection rate of 64–83% by age 13 months has been reported for countries including the United States, United Kingdom, Japan and Taiwan.[18][49] Studies have found seroprevalence varying "from approximately 39 to 80% among ethnically diverse adult populations from Tanzania, Malaysia, Thailand, and Brazil."[18] There are no significant differences among ethnic groups living in the same geographical location or between sexes. While HHV-6B is present in almost all of the world's populations, HHV-6A appears to be less frequent in Japan, North America, and Europe.[18]
Transmission
Transmission is believed to occur most frequently through the shedding of viral particles into saliva. Both HHV-6B and HHV-7 are found in human saliva, the former being at a lower frequency. Studies report varying rates of prevalence of HHV-6 in saliva (between 3–90%),[18] and have also described the salivary glands as an in vivo reservoir for HHV-6. The virus infects the salivary glands, establishes latency, and periodically reactivates to spread infection to other hosts.[28]
Vertical transmission has also been described, and occurs in approximately 1% of births in the United States.[7][50] This form is easily identifiable as the viral genome is contained within every cell of an infected individual.
Diagnosis
The diagnosis of HHV-6 infection is performed by both serologic and direct methods. The most prominent technique is the quantification of viral DNA in blood, other body fluids, and organs by means of real-time PCR.[51]
Clinical significance
The classical presentation of primary HHV-6b infection is as exanthema subitum (ES) or "roseola", featuring a high temperature lasting 3 to 5 days followed by a rash on the torso, neck, or face and sometimes febrile convulsions, however, the symptoms are not always present together. However, one study (1997) indicated that a rash is not a distinguishing feature of HHV-6 infection, with rates similar to non-HHV-6 infections (10–20% of febrile children in both groups). HHV-6 infections more frequently present with high temperatures (over 40C), at a rate of around two thirds compared to less than half in the non-HHV-6 patients. Similarly significant differences were seen in malaise, irritability, and tympanic membrane inflammation.[18]
Primary infection in adults tend to be more severe.[18]
Diagnosis for the virus, particularly HHV-6B, is vital for the patient because of the infection's adverse effects. Symptoms that point to this infection, such as rashes, go unnoticed in patients that receive antibiotics because they can be misinterpreted as a side-effect of the medicine.[18] In addition to exanthema subitum HHV-6B is known to be associated with the hepatitis, febrile convulsions, and encephalitis.
The virus periodically re-activates from its latent state, with HHV-6 DNA being detectable in 20–25% of healthy adults in the United States. In the immunocompetent setting, these re-activations are often asymptomatic, but in immunosuppressed individuals there can be serious complications. HHV-6 re-activation causes severe disease in transplant recipients and can lead to graft rejection, often in consort with other betaherpesviridae. Likewise in HIV/AIDS, HHV-6 re-activations cause disseminated infections leading to end organ disease and death. Although up to 100% of the population are exposed (seropositive) to HHV-6, most by 3 years of age, there are rare cases of primary infections in adults. In the United States, these have been linked more with HHV-6a, which is thought to be more pathogenic and more neurotropic and has been linked to several central nervous system-related disorders.
Multiple sclerosis (MS) is an autoimmune and inflammatory disorder of the nervous system that results in demyelination of axons in the brain and spinal cord. The first study to specifically investigate HHV-6-related demyelination appeared in the literature during 1996, when a previously healthy 19-month-old child developed acute encephalopathy. Levels of myelin basic protein were elevated in his cerebrospinal fluid, suggesting that demyelination was occurring.[56] This link was almost forgotten, until four years later when an MS-related study was published showing an HHV-6 prevalence of 90% among demyelinated brain tissues. In comparison, a mere 13% of disease-free brain tissues possessed the virus.[57]
The molecular mimicry hypothesis, in which T cells are essentially confusing an HHV-6 viral protein with myelin basic protein, first appeared around this time. Early on in the development of this hypothesis (2002), Italian researchers used the HHV-6a variant along with bovine myelin basic protein to generate cross-reactive T cell lines. These were compared to the T cells of individuals with MS as well as those of controls, and no significant difference was found between the two. Their early research suggested that molecular mimicry may not be a mechanism that is involved in MS.[58]
Several similar studies followed. A study from October 2014 supported the role of long-term HHV-6 infection with demyelination in progressive neurological diseases.[59]
Chronic fatigue syndrome
Chronic fatigue syndrome (CFS) is a debilitating illness,[60] the cause of which is unknown. Patients with CFS have abnormal neurological, immunological, and metabolic findings.
For many, but not all, patients who meet criteria for CFS, the illness begins with an acute, infectious-like syndrome. Cases of CFS can follow well-documented infections with several infectious agents.[61] A study of 259 patients with a "CFS-like" illness published shortly after HHV-6 was discovered used primary lymphocyte cultures to identify people with active replication of HHV-6. Such active replication was found in 70% of the patients vs. 20% of the control subjects ().[62] The question raised but not answered by this study was whether the illness caused subtle immune deficiency that led to reactivation of HHV-6, or whether reactivation of HHV-6 led to the symptoms of the illness.
Subsequent studies employing only serological techniques that do not distinguish active from latent infection have produced mixed results: most, but not all, have found an association between CFS and HHV-6 infection.[61][63][64]
Other studies have employed assays that can detect active infection: primary cell culture, PCR of serum or plasma, or IgM early antigen antibody assays. The majority of these studies have shown an association between CFS and active HHV-6 infection,[63][65][66][67][68][69] although a few have not.[64][70]
In summary, active infection with HHV-6 is present in a substantial fraction of patients with CFS. Moreover, HHV-6 is known to infect cells of the nervous system and immune system, organ systems with demonstrable abnormalities in CFS. Despite this association, it remains unproven that reactivated HHV-6 infection is a cause of CFS.
Hashimoto's thyroiditis
Hashimoto's thyroiditis is the most common thyroid disease and is characterized by abundant lymphocyte infiltrate and thyroid impairment. Recent research suggests a potential role for HHV-6 (possibly variant A) in the development or triggering of Hashimoto's thyroiditis.[71]
Pregnancy
The role of HHV-6 during pregnancy leading to inflammation in the amniotic cavity has been studied.[72]
Infertility
HHV-6A DNA was found in the endometrium of almost half of a group of infertile women, but in none of the fertile control group. Natural killer cells specific for HHV-6A, and high uterine levels of certain cytokines, were also found in the endometrium of the infertile women positive for HHV-6A. The authors suggest that HHV-6A may prove to be an important factor in female infertility.[73]
Cancer
Many human oncogenic viruses have been identified. For instance, HHV-8 is linked to Kaposi's sarcoma,[74] the Epstein–Barr virus to Burkitt's lymphoma, and HPV to cervical cancer. In fact, the World Health Organization estimated (2002) that 17.8% of human cancers were caused by infection.[75] The typical methods whereby viruses initiate oncogenesis involve suppressing the host's immune system, causing inflammation, or altering genes.
HHV-6 has been detected in lymphomas, leukemias, cervical cancers, and brain tumors.[19] Various medulloblastoma cell lines as well as the cells of other brain tumors have been demonstrated to express the CD46 receptor. Viral DNA has also been identified in many other non-pathological brain tissues, but the levels are lower.[19]
The human P53 protein functions as a tumor suppressor. Individuals who do not properly produce this protein experience a higher incidence of cancer, a phenomenon known as Li-Fraumeni syndrome. One of HHV-6's gene products, the U14 protein, binds P53 and incorporates it into virions. Another gene product, the ORF-1 protein, can also bind and inactivate P53. Cells expressing the ORF-1 gene have even been shown to produce fibrosarcomas when injected into mice.[19]
Another product of HHV-6, the immediate early protein U95, has been shown to bind nuclear factor-kappa B. Deregulation of this factor is associated with cancer.[19]
Optic neuritis
HHV-6 induced ocular inflammation has been reported three times. All three were reported in elderly individuals, two during 2007 and one during 2011. The first two were reported in Japan and France, the most recent one in Japan.[76][77][78]
These were believed to have occurred as a result of a reactivation, as anti-HHV-6 IgM antibody levels were low.[78]
Temporal lobe epilepsy
Epilepsy of the mesial temporal lobe is associated with HHV-6 infection. Within this region of the brain exists three structures: the amygdala, hippocampus, and parahippocampal gyrus. Mesial temporal lobe epilepsy (MTLE) is the most common form of chronic epilepsy and its underlying mechanism is not fully understood.[79]
Researchers consistently report having found HHV-6 DNA in tissues that were removed from patients with MTLE. Studies have demonstrated a tendency for HHV-6 to aggregate in the temporal lobe,[80] with the highest concentrations in astrocytes of the hippocampus.[79]
However, one group of researchers ultimately concluded that HHV-6 may not be involved in MTLE related to mesial temporal sclerosis.[81]
Liver failure
The virus is a common cause of liver dysfunction and acute liver failure in liver transplant recipients, and has recently been linked to periportal confluent necrosis. Furthermore, HHV-6 DNA is often detectable only in the biopsy tissues as DNA levels fall below the level of detection in blood in persistent cases.[82]
Treatment
There are no pharmaceuticals approved specifically for treating HHV-6 infection, although the usage of Cytomegalovirus treatments (valganciclovir, ganciclovir,[83]cidofovir, and foscarnet) have shown some success.[7] These drugs are given with the intent of inhibiting proper DNA polymerization by competing with deoxy triphosphate nucleotides[83] or specifically inactivating viral DNA polymerases.[2]
Finding a treatment can be difficult when HHV-6 reactivation occurs following transplant surgery because transplant medications include immunosuppressants.[84]
Related Research Articles
Cytomegalovirus (CMV) is a genus of viruses in the order Herpesvirales, in the family Herpesviridae, in the subfamily Betaherpesvirinae. Humans and other primates serve as natural hosts. The 11 species in this genus include human betaherpesvirus 5, which is the species that infects humans. Diseases associated with HHV-5 include mononucleosis and pneumonia, and congenital CMV in infants can lead to deafness and ambulatory problems.
The Epstein–Barr virus (EBV), formally called Human gammaherpesvirus 4, is one of the nine known human herpesvirus types in the herpes family, and is one of the most common viruses in humans. EBV is a double-stranded DNA virus. Epstein–Barr virus (EBV) is the first identified oncogenic virus, which establishes permanent infection in humans. EBV causes infectious mononucleosis and is also tightly linked to many malignant diseases. Various vaccine formulations underwent testing in different animals or in humans. However, none of them were able to prevent EBV infection and no vaccine has been approved to date.
Varicella zoster virus (VZV), also known as human herpesvirus 3 or Human alphaherpesvirus 3 (taxonomically), is one of nine known herpes viruses that can infect humans. It causes chickenpox (varicella) commonly affecting children and young adults, and shingles in adults but rarely in children. As a late complication of VZV infection, Ramsay Hunt syndrome type 2 may develop in rare cases. VZV infections are species-specific to humans. The virus can survive in external environments for a few hours.
Roseola, also known as sixth disease, is an infectious disease caused by certain types of human herpes viruses. Most infections occur before the age of three. Symptoms vary from absent to the classic presentation of a fever of rapid onset followed by a rash. The fever generally lasts for three to five days, while the rash is generally pink and lasts for less than three days. Complications may include febrile seizures, with serious complications being rare.
Roseolovirus is a genus of viruses in the order Herpesvirales, in the family Herpesviridae, in the subfamily Betaherpesvirinae. There are currently six species in this genus. Diseases associated with this genus include: HHV-6: sixth disease ; HHV-7: symptoms analog to the 'sixth disease'.
Kaposi's sarcoma-associated herpesvirus (KSHV) is the ninth known human herpesvirus; its formal name according to the International Committee on Taxonomy of Viruses (ICTV) is Human gammaherpesvirus 8, or HHV-8 in short. Like other herpesviruses, its informal names are used interchangeably with its formal ICTV name. This virus causes Kaposi's sarcoma, a cancer commonly occurring in AIDS patients, as well as primary effusion lymphoma, HHV-8-associated multicentric Castleman's disease and KSHV inflammatory cytokine syndrome. It is one of seven currently known human cancer viruses, or oncoviruses. Even after many years since the discovery of KSHV/HHV8, there is no known cure for KSHV associated tumorigenesis.
Virus latency is the ability of a pathogenic virus to lie dormant within a cell, denoted as the lysogenic part of the viral life cycle. A latent viral infection is a type of persistent viral infection which is distinguished from a chronic viral infection. Latency is the phase in certain viruses' life cycles in which, after initial infection, proliferation of virus particles ceases. However, the viral genome is not eradicated. The virus can reactivate and begin producing large amounts of viral progeny without the host becoming reinfected by new outside virus, and stays within the host indefinitely.
HHV Latency Associated Transcript is a length of RNA which accumulates in cells hosting long-term, or latent, Human Herpes Virus (HHV) infections. The LAT RNA is produced by genetic transcription from a certain region of the viral DNA. LAT regulates the viral genome and interferes with the normal activities of the infected host cell.
Herpesviridae is a large family of DNA viruses that cause infections and certain diseases in animals, including humans. The members of this family are also known as herpesviruses. The family name is derived from the Greek word ἕρπειν, referring to spreading cutaneous lesions, usually involving blisters, seen in flares of herpes simplex 1, herpes simplex 2 and herpes zoster (shingles). In 1971, the International Committee on the Taxonomy of Viruses (ICTV) established Herpesvirus as a genus with 23 viruses among four groups. As of 2020, 115 species are recognized, all but one of which are in one of the three subfamilies. Herpesviruses can cause both latent and lytic infections.
Herpes simplex virus1 and 2, also known by their taxonomic names Human alphaherpesvirus 1 and Human alphaherpesvirus 2, are two members of the human Herpesviridae family, a set of viruses that produce viral infections in the majority of humans. Both HSV-1 and HSV-2 are very common and contagious. They can be spread when an infected person begins shedding the virus.
Gammaherpesvirinae is a subfamily of viruses in the order Herpesvirales and in the family Herpesviridae. Viruses in Gammaherpesvirinae are distinguished by reproducing at a more variable rate than other subfamilies of Herpesviridae. Mammals serve as natural hosts. There are 43 species in this subfamily, divided among 7 genera with three species unassigned to a genus. Diseases associated with this subfamily include: HHV-4: infectious mononucleosis. HHV-8: Kaposi's sarcoma.
Betaherpesvirinae is a subfamily of viruses in the order Herpesvirales and in the family Herpesviridae. Mammals serve as natural hosts. There are 26 species in this subfamily, divided among 5 genera. Diseases associated with this subfamily include: human cytomegalovirus (HHV-5): congenital CMV infection; HHV-6: 'sixth disease' ; HHV-7: symptoms analogous to the 'sixth disease'.
Human betaherpesvirus 5, also called human cytomegalovirus (HCMV,HHV-5), is a species of virus in the genus Cytomegalovirus, which in turn is a member of the viral family known as Herpesviridae or herpesviruses. It is also commonly called CMV. Within Herpesviridae, HCMV belongs to the Betaherpesvirinae subfamily, which also includes cytomegaloviruses from other mammals. CMV is a double-stranded DNA virus.
Human betaherpesvirus 7 (HHV-7) is one of nine known members of the Herpesviridae family that infects humans. HHV-7 is a member of Betaherpesvirinae, a subfamily of the Herpesviridae that also includes HHV-6 and Cytomegalovirus. HHV-7 often acts together with HHV-6, and the viruses together are sometimes referred to by their genus, Roseolovirus. HHV-7 was first isolated in 1990 from CD4+ T cells taken from peripheral blood lymphocytes.
Neurovirology is an interdisciplinary field which represents a melding of clinical neuroscience, virology, immunology, and molecular biology. The main focus of the field is to study viruses capable of infecting the nervous system. In addition to this, the field studies the use of viruses to trace neuroanatomical pathways, for gene therapy, and to eliminate detrimental populations of neural cells.
Dharam Vir Ablashi was an American biomedical researcher born in Lahore. He is best known for his co-discovery of Human herpesvirus 6 (HHV-6), an immunosuppressive and neurotropic virus that can cause encephalitis and seizures during a primary infection or when reactivated from latency in immunosuppressed patients.
Konstance K. Knox is an American virologist and entrepreneur who founded Coppe Laboratories, Viracor, the Wisconsin Viral Research Group (WVRG) and Viracor's Institute for Viral Pathogenesis (IVP). In 2000, Viracor became the first biotechnology company in the United States to correlate multiple sclerosis with human herpesvirus 6 (HHV-6).
Human herpes viruses, also known as HHVs, are part of a family of DNA viruses that cause several diseases in humans. One of the most notable functions of this virus family is their ability to enter a latent phase and lay dormant within animals for extended periods of time. The mechanism that controls this is very complex because expression of viral proteins during latency is decreased a great deal, meaning that the virus must have transcription of its genes repressed. There are many factors and mechanisms that control this process and epigenetics is one way this is accomplished. Epigenetics refers to persistent changes in expression patterns that are not caused by changes to the DNA sequence. This happens through mechanisms such as methylation and acetylation of histones, DNA methylation, and non-coding RNAs (ncRNA). Altering the acetylation of histones creates changes in expression by changing the binding affinity of histones to DNA, making it harder or easier for transcription machinery to access the DNA. Methyl and acetyl groups can also act as binding sites for transcription factors and enzymes that further modify histones or alter the DNA itself.
Human betaherpesvirus 6A (HHV-6A) is a species of virus in the genus Roseolovirus, subfamily Betaherpesvirinae, family Herpesviridae, and order Herpesvirales.
Human betaherpesvirus 6B (HHV-6B) is a species of virus in the genus Roseolovirus, subfamily Betaherpesvirinae, family Herpesviridae, and order Herpesvirales.
↑ De Bolle, L.; Van Loon, J.; De Clercq, E.; Naesens, L. (2005). "Quantitative analysis of human herpesvirus 6 cell tropism". Journal of Medical Virology. 75 (1): 76–85. doi:10.1002/jmv.20240. PMID15543581. S2CID31720143.
↑ Álvarez-Lafuente, Roberto; García-Montojo, Marta; De Las Heras, Virginia; Bartolomé, Manuel; Arroyo, Rafael (2006). "Clinical parameters and HHV-6 active replication in relapsing—remitting multiple sclerosis patients". Journal of Clinical Virology. 37: S24–6. doi:10.1016/S1386-6532(06)70007-5. PMID17276363.
↑ Anderson, L. (1988). "Human Retrovirus Family: Cancer, Central Nervous System Disease, and AIDS". JNCI Journal of the National Cancer Institute. 80 (13): 987–9. doi:10.1093/jnci/80.13.987. PMID2842514.
1 2 Salahuddin, S.; Ablashi, D.; Markham, P.; Josephs, S.; Sturzenegger, S; Kaplan, M; Halligan, G; Biberfeld, P; etal. (1986). "Isolation of a new virus, HBLV, in patients with lymphoproliferative disorders". Science. 234 (4776): 596–601. Bibcode:1986Sci...234..596Z. doi:10.1126/science.2876520. PMID2876520.
↑ Fenner, F. (1971). "The nomenclature and classification of viruses the International Committee on Nomenclature of Viruses". Virology. 46 (3): 979–980. doi:10.1016/0042-6822(71)90102-4. PMID18619371.
↑ J Exp Med. 1995 Apr 1;181(4):1303–10. Infection of gamma/delta T lymphocytes by human herpesvirus 6: transcriptional induction of CD4 and susceptibility to HIV infection. Lusso P, Garzino-Demo A, Crowley RW, Malnati MS.
1 2 Liedtke, W.; Opalka, B.; Zimmermann, C.W.; Lignitz, E. (1993). "Age distribution of latent herpes simplex virus 1 and varicella-zoster virus genome in human nervous tissue". Journal of the Neurological Sciences. 116 (1): 6–11. doi:10.1016/0022-510X(93)90082-A. PMID8389816. S2CID32218673.
↑ Lusso, Paolo; De Maria, Andrea; Malnati, Mauro; Lori, Franco; Derocco, Susan E.; Baseler, Michael; Gallo, Robert C. (1991). "Induction of CD4 and susceptibility to HIV-1 infection in human CD8+ T lymphocytes by human herpesvirus 6". Nature. 349 (6309): 533–5. Bibcode:1991Natur.349..533L. doi:10.1038/349533a0. PMID1846951. S2CID4307070.
↑ Arena, A; Liberto, MC; Capozza, AB; Focà, A (1997). "Productive HHV-6 infection in differentiated U937 cells: Role of TNF alpha in regulation of HHV-6". The New Microbiologica. 20 (1): 13–20. PMID9037664.
↑ Inagi, Reiko; Guntapong, Ratigorn; Nakao, Masayuki; Ishino, Yoshizumi; Kawanishi, Kazunobu; Isegawa, Yuji; Yamanishi, Koichi (1996). "Human herpesvirus 6 induces IL-8 gene expression in human hepatoma cell line, Hep G2". Journal of Medical Virology. 49 (1): 34–40. doi:10.1002/(SICI)1096-9071(199605)49:1<34::AID-JMV6>3.0.CO;2-L. PMID8732869. S2CID34843836.
↑ Araujo, A.; Pagnier, A.; Frange, P.; Wroblewski, I.; Stasia, M.-J.; Morand, P.; Plantaz, D. (2011). "Syndrome d'activation lymphohistiocytaire associé à une infection à Burkholderia cepacia complex chez un nourrisson révélant une granulomatose septique et une intégration génomique du virus HHV-6" [Lymphohistiocytic activation syndrome and Burkholderia cepacia complex infection in a child revealing chronic granulomatous disease and chromosomal integration of the HHV-6 genome]. Archives de Pédiatrie (in French). 18 (4): 416–9. doi:10.1016/j.arcped.2011.01.006. PMID21397473.
↑ Komaroff, Anthony L. (2006). "Is human herpesvirus-6 a trigger for chronic fatigue syndrome?". Journal of Clinical Virology. 37: S39–46. doi:10.1016/S1386-6532(06)70010-5. PMID17276367.
↑ Pietilainen-Nicklen, J.; Virtanen, J (2014). "HHV-6-positivity in diseases with demyelination". Journal of Clinical Virology. 61 (2): 216–9. doi:10.1016/j.jcv.2014.07.006. PMID25088617.
↑ Fukuda, K; Straus, SE; Hickie, I; Sharpe, MC; Dobbins, JG; Komaroff, A (1994). "The chronic fatigue syndrome: A comprehensive approach to its definition and study. International Chronic Fatigue Syndrome Study Group". Annals of Internal Medicine. 121 (12): 953–9. doi:10.7326/0003-4819-121-12-199412150-00009. PMID7978722. S2CID510735.
↑ Nicolson, G. L.; Gan, R.; Haier, J. (2003). "Multiple co-infections (Mycoplasma, Chlamydia, human herpes virus-6) in blood of chronic fatigue syndrome patients: Association with signs and symptoms". APMIS. 111 (5): 557–66. doi:10.1034/j.1600-0463.2003.1110504.x. PMID12887507. S2CID20406603.
↑ Patnaik, M.; Komaroff, A. L.; Conley, E.; Ojo-Amaize, E. A.; Peter, J. B. (1995). "Prevalence of IgM Antibodies to Human Herpesvirus 6 Early Antigen (p41/38) in Patients with Chronic Fatigue Syndrome". Journal of Infectious Diseases. 172 (5): 1364–7. doi:10.1093/infdis/172.5.1364. PMID7594679.
↑ Secchiero, P.; Carrigan, D. R.; Asano, Y.; Benedetti, L.; Crowley, R. W.; Komaroff, A. L.; Gallo, R. C.; Lusso, P. (1995). "Detection of Human Herpesvirus 6 in Plasma of Children with Primary Infection and Immunosuppressed Patients by Polymerase Chain Reaction". Journal of Infectious Diseases. 171 (2): 273–80. doi:10.1093/infdis/171.2.273. PMID7844362.
↑ Wagner, Mathias; Krueger, Gerhard; Ablashi, Dharam; Whitman, James (1996). "Chronic Fatigue Syndrome (CFS): A Critical Evaluation of Testing for Active Human Herpesvirus-6 (HHV-6) Infection". Journal of Chronic Fatigue Syndrome. 2 (4): 3–16. doi:10.1300/J092v02n04_02.
↑ Silasi, Michelle (March 2013). "Viral invasion of the Amniotic Cavity (VIAC) in the Midtrimester of Pregnancy". American Journal of Reproductive Immunology. 69 (3): 195–196. doi:10.1111/aji.12073. PMID23384234. S2CID747338.
↑ Oberacher-Velten, Isabel M.; Jonas, Jost B.; Jünemann, Anselm; Schmidt, Barbara (2004). "Bilateral optic neuropathy and unilateral tonic pupil associated with acute human herpesvirus 6 infection: A case report". Graefe's Archive for Clinical and Experimental Ophthalmology. 243 (2): 175–7. doi:10.1007/s00417-004-0986-8. PMID15742213. S2CID12794574.
↑ Maslin, Jérôme; Bigaillon, Christine; Froussard, Françoise; Enouf, Vincent; Nicand, Elisabeth (2007). "Acute bilateral uveitis associated with an active human herpesvirus-6 infection". Journal of Infection. 54 (4): e237–40. doi:10.1016/j.jinf.2006.12.012. PMID17303245.
1 2 Ogata, Nahoko; Koike, Naoko; Yoshikawa, Tadanobu; Takahashi, Kanji (2011). "Human herpesvirus 6-associated uveitis with optic neuritis diagnosed by multiplex PCR". Japanese Journal of Ophthalmology. 55 (5): 502–5. doi:10.1007/s10384-011-0069-4. PMID21814813. S2CID189770831.
1 2 Li, Jin-Mei; Lei, Ding; Peng, Fang; Zeng, Yi-Jun; Li, Liang; Xia, Zeng-Liang; Xia, Xiao-Qiang; Zhou, Dong (2011). "Detection of human herpes virus 6B in patients with mesial temporal lobe epilepsy in West China and the possible association with elevated NF-κB expression". Epilepsy Research. 94 (1–2): 1–9. doi:10.1016/j.eplepsyres.2010.11.001. PMID21256714. S2CID23341675.
↑ Noguchi, T.; Yoshiura, T.; Hiwatashi, A.; Togao, O.; Yamashita, K.; Nagao, E.; Uchino, A.; Hasuo, K.; etal. (2010). "CT and MRI Findings of Human Herpesvirus 6-Associated Encephalopathy: Comparison with Findings of Herpes Simplex Virus Encephalitis". American Journal of Roentgenology. 194 (3): 754–60. doi:10.2214/AJR.09.2548. PMID20173155.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.