
Agnosia is the inability to process sensory information. Often there is a loss of ability to recognize objects, persons, sounds, shapes, or smells while the specific sense is not defective nor is there any significant memory loss. It is usually associated with brain injury or neurological illness, particularly after damage to the occipitotemporal border, which is part of the ventral stream. Agnosia only affects a single modality, such as vision or hearing. More recently, a top-down interruption is considered to cause the disturbance of handling perceptual information.

The temporal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The temporal lobe is located beneath the lateral fissure on both cerebral hemispheres of the mammalian brain.

Facial perception is an individual's understanding and interpretation of the face. Here, perception implies the presence of consciousness and hence excludes automated facial recognition systems. Although facial recognition is found in other species, this article focuses on facial perception in humans.

Prosopagnosia, also known as face blindness, is a cognitive disorder of face perception in which the ability to recognize familiar faces, including one's own face (self-recognition), is impaired, while other aspects of visual processing and intellectual functioning remain intact. The term originally referred to a condition following acute brain damage, but a congenital or developmental form of the disorder also exists, with a prevalence of 2.5%. The brain area usually associated with prosopagnosia is the fusiform gyrus, which activates specifically in response to faces. The functionality of the fusiform gyrus allows most people to recognize faces in more detail than they do similarly complex inanimate objects. For those with prosopagnosia, the method for recognizing faces depends on the less sensitive object-recognition system. The right hemisphere fusiform gyrus is more often involved in familiar face recognition than the left. It remains unclear whether the fusiform gyrus is specific for the recognition of human faces or if it is also involved in highly trained visual stimuli. Prosopoagnosic patients are under normal conditions able to recognize facial expressions and emotions.

The fusiform gyrus, also known as the lateral occipitotemporal gyrus,is part of the temporal lobe and occipital lobe in Brodmann area 37. The fusiform gyrus is located between the lingual gyrus and parahippocampal gyrus above, and the inferior temporal gyrus below. Though the functionality of the fusiform gyrus is not fully understood, it has been linked with various neural pathways related to recognition. Additionally, it has been linked to various neurological phenomena such as synesthesia, dyslexia, and prosopagnosia.
Visual processing is a term that is used to refer to the brain's ability to use and interpret visual information from the world around us. The process of converting light energy into a meaningful image is a complex process that is facilitated by numerous brain structures and higher level cognitive processes. On an anatomical level, light energy first enters the eye through the cornea, where the light is bent. After passing through the cornea, light passes through the pupil and then lens of the eye, where it is bent to a greater degree and focused upon the retina. The retina is where a group of light-sensing cells, called photoreceptors are located. There are two types of photoreceptors: rods and cones. Rods are sensitive to dim light and cones are better able to transduce bright light. Photoreceptors connect to bipolar cells, which induce action potentials in retinal ganglion cells. These retinal ganglion cells form a bundle at the optic disc, which is a part of the optic nerve. The two optic nerves from each eye meet at the optic chiasm, where nerve fibers from each nasal retina cross which results in the right half of each eye's visual field being represented in the left hemisphere and the left half of each eye's visual fields being represented in the right hemisphere. The optic tract then diverges into two visual pathways, the geniculostriate pathway and the tectopulvinar pathway, which send visual information to the visual cortex of the occipital lobe for higher level processing.

Visual memory describes the relationship between perceptual processing and the encoding, storage and retrieval of the resulting neural representations. Visual memory occurs over a broad time range spanning from eye movements to years in order to visually navigate to a previously visited location. Visual memory is a form of memory which preserves some characteristics of our senses pertaining to visual experience. We are able to place in memory visual information which resembles objects, places, animals or people in a mental image. The experience of visual memory is also referred to as the mind's eye through which we can retrieve from our memory a mental image of original objects, places, animals or people. Visual memory is one of several cognitive systems, which are all interconnected parts that combine to form the human memory. Types of palinopsia, the persistence or recurrence of a visual image after the stimulus has been removed, is a dysfunction of visual memory.

In psycholinguistics, language processing refers to the way humans use words to communicate ideas and feelings, and how such communications are processed and understood. Language processing is considered to be a uniquely human ability that is not produced with the same grammatical understanding or systematicity in even human's closest primate relatives.

Associative visual agnosia is a form of visual agnosia. It is an impairment in recognition or assigning meaning to a stimulus that is accurately perceived and not associated with a generalized deficit in intelligence, memory, language or attention. The disorder appears to be very uncommon in a "pure" or uncomplicated form and is usually accompanied by other complex neuropsychological problems due to the nature of the etiology. Affected individuals can accurately distinguish the object, as demonstrated by the ability to draw a picture of it or categorize accurately, yet they are unable to identify the object, its features or its functions.
The two-streams hypothesis is a model of the neural processing of vision as well as hearing. The hypothesis, given its initial characterisation in a paper by David Milner and Melvyn A. Goodale in 1992, argues that humans possess two distinct visual systems. Recently there seems to be evidence of two distinct auditory systems as well. As visual information exits the occipital lobe, and as sound leaves the phonological network, it follows two main pathways, or "streams". The ventral stream leads to the temporal lobe, which is involved with object and visual identification and recognition. The dorsal stream leads to the parietal lobe, which is involved with processing the object's spatial location relative to the viewer and with speech repetition.
Visual agnosia is an impairment in recognition of visually presented objects. It is not due to a deficit in vision, language, memory, or intellect. While cortical blindness results from lesions to primary visual cortex, visual agnosia is often due to damage to more anterior cortex such as the posterior occipital and/or temporal lobe(s) in the brain.[2] There are two types of visual agnosia: apperceptive agnosia and associative agnosia.

The inferior temporal gyrus is one of three gyri of the temporal lobe and is located below the middle temporal gyrus, connected behind with the inferior occipital gyrus; it also extends around the infero-lateral border on to the inferior surface of the temporal lobe, where it is limited by the inferior sulcus. This region is one of the higher levels of the ventral stream of visual processing, associated with the representation of objects, places, faces, and colors. It may also be involved in face perception, and in the recognition of numbers and words.

The colour centre is a region in the brain primarily responsible for visual perception and cortical processing of colour signals received by the eye, which ultimately results in colour vision. The colour centre in humans is thought to be located in the ventral occipital lobe as part of the visual system, in addition to other areas responsible for recognizing and processing specific visual stimuli, such as faces, words, and objects. Many functional magnetic resonance imaging (fMRI) studies in both humans and macaque monkeys have shown colour stimuli to activate multiple areas in the brain, including the fusiform gyrus and the lingual gyrus. These areas, as well as others identified as having a role in colour vision processing, are collectively labelled visual area 4 (V4). The exact mechanisms, location, and function of V4 are still being investigated.

The fusiform face area is a part of the human visual system that is specialized for facial recognition. It is located in the inferior temporal cortex (IT), in the fusiform gyrus.

The inferior longitudinal fasciculus (ILF) is traditionally considered one of the major occipitotemporal association tracts. It is the white matter backbone of the ventral visual stream. It connects the ventral surface of the anterior temporal lobe and the extrastriate cortex of the occipital lobe, running along the lateral and inferior wall of the lateral ventricle.
Apperceptive agnosia is a failure in recognition that is due to a failure of perception. In contrast, associative agnosia is a type of agnosia where perception occurs but recognition still does not occur. When referring to apperceptive agnosia, visual and object agnosia are most commonly discussed; this occurs because apperceptive agnosia is most likely to present visual impairments. However, in addition to visual apperceptive agnosia there are also cases of apperceptive agnosia in other sensory areas.
Recognition memory, a subcategory of explicit memory, is the ability to recognize previously encountered events, objects, or people. When the previously experienced event is reexperienced, this environmental content is matched to stored memory representations, eliciting matching signals. As first established by psychology experiments in the 1970s, recognition memory for pictures is quite remarkable: humans can remember thousands of images at high accuracy after seeing each only once and only for a few seconds.
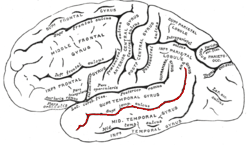
In the human brain, the superior temporal sulcus (STS) is the sulcus separating the superior temporal gyrus from the middle temporal gyrus in the temporal lobe of the brain. A sulcus is a deep groove that curves into the largest part of the brain, the cerebrum, and a gyrus is a ridge that curves outward of the cerebrum.
Form perception is the recognition of visual elements of objects, specifically those to do with shapes, patterns and previously identified important characteristics. An object is perceived by the retina as a two-dimensional image, but the image can vary for the same object in terms of the context with which it is viewed, the apparent size of the object, the angle from which it is viewed, how illuminated it is, as well as where it resides in the field of vision. Despite the fact that each instance of observing an object leads to a unique retinal response pattern, the visual processing in the brain is capable of recognizing these experiences as analogous, allowing invariant object recognition. Visual processing occurs in a hierarchy with the lowest levels recognizing lines and contours, and slightly higher levels performing tasks such as completing boundaries and recognizing contour combinations. The highest levels integrate the perceived information to recognize an entire object. Essentially object recognition is the ability to assign labels to objects in order to categorize and identify them, thus distinguishing one object from another. During visual processing information is not created, but rather reformatted in a way that draws out the most detailed information of the stimulus.
Patient DF is a woman with visual apperceptive agnosia who has been studied extensively due to the implications of her behavior for the two streams theory of visual perception. Though her vision remains intact, she has trouble visually locating and identifying objects. Her agnosia is thought to be caused by a bilateral lesion to her lateral occipital cortex, an area thought by dual-stream proponents to be the ventral "object recognition" stream. Despite being unable to identify or recognize objects, DF can still use visual input to guide her action.

